
The Genesis of Pathogenic E. coli
Abstract
Even though some strains are pathogenic, most E. coli strains still show evidence of being one of God’s “very good” creations.
Keywords: E. coli, Escherichia coli, Shigella, mutualism, genesis of germs, interwoven complexity, biomatrix, Theodor Escherich, history of intestinal diseases
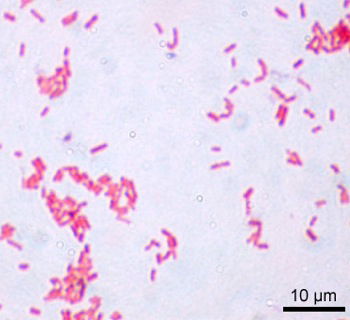
Fig. 1. E. coli Gram stain (Wiki commons image). E. coli are Gram-negative bacteria, thus red or pink colored. The red color is due to a counterstain, called safranin.
Escherichia coli is frequently in the news (Fig. 1). E. coli often gets “bad press” for contaminating drinking water or causing a food-borne infection (via hamburgers, apple juice, spinach, or other foods). Recently a new strain (E. coli O145) has been implicated in contaminating lettuce in the U.S., while another strain (E. coli O157:H7) is apparently in tons of beef and other foods. Pathogenic (disease-causing) E. coli is becoming so common in foods that the government is likely to “beef up” its regulation on the food industry (Dininny 2010). Even though some strains are pathogenic, most E. coli strains still show evidence of being one of God’s “very good” creations. E. coli is also a common experimental organism (a laboratory “pet”) of biologists, and is valuable for studying genetics and variation in living things.
Newspapers, biology texts and the popular media increasingly discuss “evolution in action.” Evidence to support this concept includes emerging diseases, antibiotic resistance, and changes in characteristics of bacteria, especially the work of Richard Lenski’s lab. Lenski and his coworkers have shown that bacteria can change rapidly in phenotype (outward appearance) and evolutionists have seized upon this rapid phenotypic change as alleged powerful evidence for Darwinian evolution. Thus, E. coli and “evolution in action” is an important two-fold issue involving empirical (or observational/operational) science and its relationship to the theoretical (or historical) origins issue.
Carl Zimmer (2008), in his book, Microcosm: E. coli and the New Science of Life, uses emerging diseases caused by E. coli to bolster his arguments for molecules-to-man evolution. Zimmer’s book popularizes the work of Lenski on emerging (i.e., newly appearing) diseases and increases the appeal of his research as seen in news magazines, general biology and microbiology texts, and at many scientific conferences. Zimmer states that scientists are investigating phenotypic changes in E. coli to demonstrate their ability to undergo “rapid bursts of evolution” (p. 97). Many biologists argue that the wide variation in pathogenicity found in E. coli as support for Darwinian evolution. The typical explanation for the origin of E. coli is that it has been around for billions of years and man only a few millions years. For the mutualistic relationship to have begun between man and microbe, animals and E. coli had to co-evolve in both E. coli and man. The human body had to allow a bacteria to pass through the stomach and reach the intestine (a rapid turnover rate) to become one of the most successful bacteria on the planet (i.e., it's found in every mammal known and even extends to fish).
Increasingly reoccurring themes (examples) are being declared about pathogenic E. coli and its “evolution in action”: primarily how new and emerging diseases arise. The origins of new diseases within the same species are really examples of variation and adaptation. We seek to provide an “answer” and alternative to the Darwinian paradigm through investigation of the origin of E. coli and its role in examples of “evolution in action.” The purpose of this article is to discuss the possible origin of E. coli from the time of Creation; and its modification since the Edenic Curse. The specific objectives of this article are to provide reasonable explanations for: (1) the origin and purpose of E. coli in the human body; (2) the genesis of new pathogenic E. coli strains; and (3) a reasonable alternative to evolution in regard to past changes in E. coli and similar bacteria. Finally, we discuss how E. coli fits into the historic, biblical worldview stages of Creation, Curse, Corruption, and Contagion.3 It will integrate the topics of modification and displacement for emerging diseases.
The Origin of E. coli
We read in Genesis 1 that God made everything “very good.” If everything God made was good, where did pathogenic E. coli come from? What is the origin of this frightening germ? Where does it fit into the creation account? Was it created along with the rest of life, during the first week of Creation, or later, after the Fall of man? Is E. coli a result of the “Curse”? These and other questions have been asked by professional creation microbiologists (i.e., Francis 2003; Gillen 2007; Gillen 2008). Their answers may surprise you.

Fig. 2. Theodor Escherich (Wiki commons image). Theodor Escherich (1857–1911) was a pediatrician and discovered E. coli in 1885.
The coliform (rod-shaped, intestinal bacterial) “kind” (including Escherichia and Shigella species) is a rather hardy, Gram-negative bacterial type (i.e. they do not become colored if put into a purple Gram stain, and eventually stain red with a counterstain) that is especially resistant to acid and adapts to a variety of oxygen conditions. These bacteria usually occur as single unattached rods (bacillus) (Fig. 1). The tiny rods measure about 2 µm (micrometers) in length and 0.5 µm in diameter. Coliform bacteria grow at a variety of temperatures and oxygen conditions, but seem to do especially well at normal human body temperature (37 °C), and can grow in aqueous environments (Talaro 2008).
Microbiology research based on the creation paradigm appears to provide answers to some puzzling questions regarding the E. coli “kind” (including Shigella spp.). The genera Shigella and Escherichia probably are from the same original Biblical kind, based on their similar appearance, gut habitat, genome sequence, and biochemical properties. We could call the first set of originally created coliforms, the E. coli prototype. For over a decade (1883–1897) these bacteria were collectively called Bacterium coli, according to Theodor Escherich (1883) (Fig. 2).
We cannot be dogmatic (beyond the biblical text) regarding details of microbe origins during Creation week, but believe that Scripture and biological data suggest something about the nature of microbes in a mature but fallen creation. Diligent research by past creation biologists1 such as Leeuwenhoek, Pasteur, and Lister, were blessed by God as He revealed to them critical insight into His creation (Psalm 139:17; Gillen and Sherwin 2008). We hope and pray to give a reasonable modern analysis.
The Coliform “Kind”
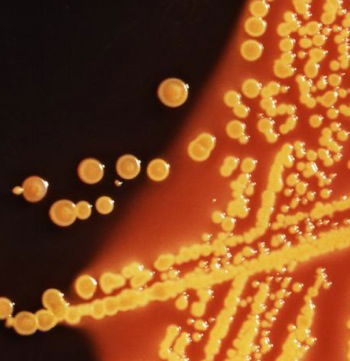
Fig. 3. E. coli colonies (CDC image # 6676). These are colonies of E. coli bacteria grown on an Hektoen enteric (HE) agar plate medium. E. coli grown on HE agar display a raised morphology, and are yellow, to orange-yellow in coloration.
Although E. coli frequently gets “bad press” for contaminating drinking water or causing a food-borne infection, most E. coli are “very good” creations, even in the present world (Fig. 3). E. coli is a normal inhabitant of the large intestine of vertebrates, including humans, and its presence is beneficial because it helps produce certain vitamins and breaks down otherwise indigestible foodstuffs. E. coli are part of our normal microflora (also called normal flora) in human intestines (Fig. 4).
Without our intestinal flora, we would not survive very well. E. coli and other enteric bacteria supply our body with vitamin K, vitamin B12, thiamin, and riboflavin, which we need for normal body functioning. These intestinal microflora also stimulate lymph node-like structures, called Peyer’s patches, which contain lymphatic tissue and provide the intestines with protection, even helping to prevent colon cancer. They maintain an intensely competitive and closed community, which makes invasion from pathogens a considerable challenge. They produce substances to prevent harmful microbes from invading. They can serve as a probiotic, i.e., living organisms that promote health. As for the benefit to E. coli, man’s colon provides them a stable nutritional home. In summary, E. coli provides humans with vitamins and body defenses as its “rent money,” and, in turn, the human colon provides it with a safe and warm home: it is a mutualistic relationship (Fig. 4).
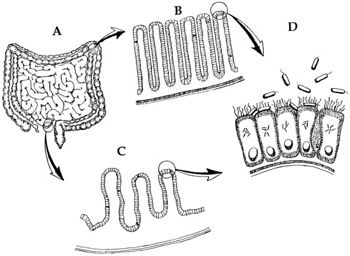
Fig. 4. Original Design. Normal flora and boundaries maintained. Most of the time, bacteria (e.g., nonpathogenic E. coli) inhabit the gastrointestinal (GI) tract mutualistically. A illustrates the small and large intestines. B is a close up of the villus lining. C is a close up of the microvillus lining, the site where nutrients are absorbed and transferred to the bloodstream. D illustrates the site where E. coli assists in the absorption of micronutrients and manufactures B-vitamins at the surface of the villi (adapted from Gillen, Sherwin and Knowles 2001, p. 107).
E. coli and Its Extension of the Interwoven Complexity Found in the Digestive System
An interwoven, adaptation package is a biological relationship in which the whole is functionally more than the sum of its individual components. We call this an extension of the interwoven complexity observed in the human body (Gillen 2006). Dr. Joseph Francis calls it the “biomatrix” of life, or “organosubstrate” theory. E. coli (Fig. 2) are the predominant bacteria in the lower intestine, and they constitute about 75 percent of all living bacteria in feces.
Indeed, the intestinal bacteria contribute to the general well-being of both microbes and people by synthesizing a number of the vitamins essential for good nutrition and breaking down various macronutrients. The human body cannot synthesize niacin to make nicotinamide adenine dinucleotide (NAD) that is necessary for energy conversion in the cell’s mitochondria. The B-vitamins, including niacin, cobalamin (B12) biotin, thiamine, and riboflavin, are necessary for normal energy levels, freedom from fatigue, and proper functioning of nerves. A prolonged deficiency in any one of these vitamins may lead to chronic fatigue and inability to lead a normal life. As for the benefit to E. coli (and other enteric bacteria), man’s colon provides them a stable nutritional home. In short, both parties benefit from this mutualistic relationship.
The mutualistic relationship between our digestive cells and these helpful microbes shows amazing cooperation and is another example of emergent properties in living things. The large intestine contains the highest numbers of resident flora because of the available moisture and nutrients in the colon. If you take away E. coli and other bacteria, the function of the digestive system is severely impaired. This cooperation among cells is similar to the interdependent parts of a mousetrap and summarized in the table below.
Table 1. Benefits of E. coli and Other Coliform Bacteria to the Human Body
| Vitamins and Enzymes | Physiological Role | Disease if Deficient |
|---|---|---|
| 1. Vitamin K | Needed in blood clotting | Hemorrhage in newborns who lack gut bacteria |
| 2. Vitamin B12 (absorbs) | Coenzyme needed in protein formation | Pernicious anemia; lack of energy |
| 3. Niacin (Vitamin B3) | Part of coenzymes in energy metabolism | Pellagra; lack of energy; fatigue |
| 4. Vitamin B-Complex (thiamine, biotin, and riboflavin) | Carbohydrate metabolism
Coenzymes in cell respiration | Beriberi, loss of appetite, fatigue, inflammation and breakdown of skin |
Pre-Fall World and the Normal Microflora of the Intestine
Where do the E. coli microbes fit into God’s “very good” creation? Most creation biologists believe that God created (Genesis 1 and 2) all bacteria as very good forms of life (Francis 2003; Gillen 2007). Even today, a vast majority of bacteria are beneficial to man and nature. Only about 5 to 10% of all bacteria are pathogenic. Many microbes live in a mutualistic relationship with other microbes, but they also reside with plants, animals, and humans. Mutualism is a type of symbiosis (Greek sym; together, bios, life). It is “normal” and critical for our body’s health to be symbiotically inhabited by microbes such as beneficial bacteria. God’s very good creation included design features involving important microbes. This internal and external beneficial colonization of the body is termed our normal microbiota.
In some cases the symbiosis is beneficial to both the body and the microorganisms; this relationship is called mutualism. Humans are “covered” outside and inside with millions of microbiotic organisms that maintain normal bodily functions and sustain life in changing environments. For example, many Staphylococcus species live on human skin. S. epidermidis alone makes up 90% of the normal flora on the skin (Gillen 2009). In a brilliant stroke, the Creator most likely wove2 together man’s body with staphylococci outside on the skin, and on the inside, enteric bacteria (like E. coli) in the intestine. In Genesis 2, we believe that man was formed fully mature with the appearance of an adult. Somewhat like an embroidered quilt, He likely made humans with bacteria on their skin, and in their intestines.
The normal flora provides clues to the pre-Fall function of bacteria (Fig. 4). The intestine provides a microbiome for many bacteria. Dr. Francis (Francis 2003) suggests that microbes are a natural healthy extension of the body and that bacteria form a biomatrix (technically an organosubstrate) on the epidermis. When God created man, most likely He made the Escherichia/Shigella kind to help maintain the digestive and immune system. Over time, minor changes took place (i.e. variation): some of the enteric bacterial strains came to occupy most of the small and large intestines. Perhaps, the original Escherichia kind diversified into several varieties according to microhabitat: some to live in the small intestine (ileum, duodenum, and jejunum), and some in specialized places of the large intestine (ascending, transverse, and descending colon), and rectum.
Most likely, Escherichia was originally designed to live in harmony with man as a harmless member of the gut flora. It played a positive role in producing vitamins and helping with digestion with other enteric bacteria in 34 genera (~ 5000 species). It is likely this was part of their original good design. As Francis (2009) points out, the gut is a “living city” of various microbes. However, after the Edenic Fall, the Curse of the Creator caused corruption and decay in these living bacteria, including in their genetic code. Today, the term germ specifically refers to disease-causing microbes, or pathogens (Gillen 2007). All “germs” would have had their origin after the Fall (Genesis 3). The Edenic Curse would have profoundly influenced all Creation, including those viruses, bacteria, fungi, and protozoans that would later become pathogens or parasites.
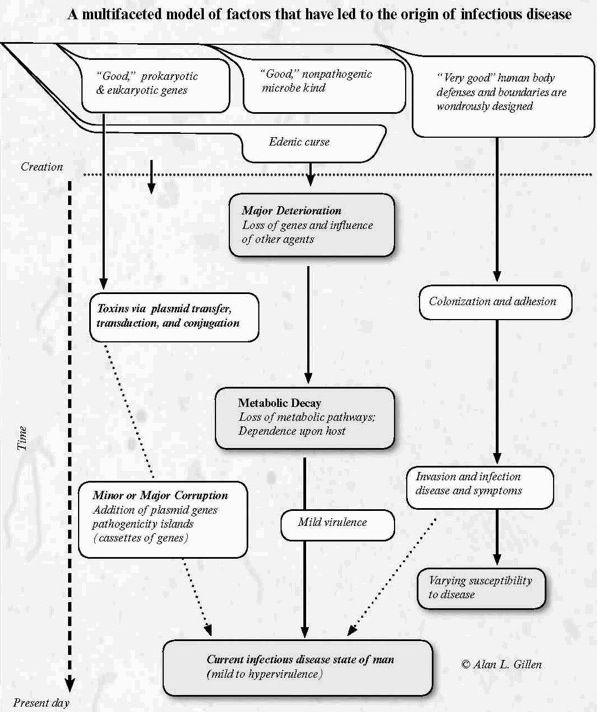
Fig. 5. Genesis of Germs summary (Genesis of Germs, Fig. 9.1, Master Books). The origin of infectious diseases is multifaceted and very complex.
Clearly, the origin of infectious disease is complex and multifaceted (Fig. 5). This topic is further explored in the book, The Genesis of Germs (Gillen 2007). It provides some understanding into the origin of such diseases. From a biblical worldview, infectious diseases and pathogenesis had become a secondary condition, a result of sin that was not what the Creator had intended. But even in this now-fallen world, God provided humans with defenses such as members of the gut’s microbiota that secrete chemicals to interfere with the growth of pathogenic microbiota (Bauman 2009).
History of Intestinal Diseases and Human Waste Disposal in Ancient Times
Fig. 6. E. coli chronic diarrhea (CDC image # 2156). Virulence factors include capsule, fimbria, endotoxins, adhesions, exotoxins, and type III secretion system.
S. dysenteriae and E. coli are likely within the same “biblical kind.” S. dysenteriae is a species of the rod-shaped bacterial genus, Shigella. (The Shigellae are known as Gram-negative, non-spore-forming, facultative anaerobic, non-motile bacteria.) Like E. coli, Shigella can cause bacillary dysentery (Fig. 6). (Dysentery is bloody diarrhea.) No one knows with certainty the origins of E. coli diarrheal disease. However, it appears that it is a very old disease. The Jews were instructed to dispose of their human waste outside the camp (Deuteronomy 23:12–13). In Deuteronomy 23:12–13, the Lord gave instructions to the Israelites for a sewage system that is not dissimilar to what we use today:
“Also you shall have a place outside the camp, where you may go out; and you shall have an implement among your equipment, and when you sit down outside, you shall dig with it and turn and cover your refuse” (Deuteronomy 23:12–13).
The Creator told His people that they should designate a place outside their camp and relieve themselves; when they go they should take a shovel; then, dig a hole and cover their human waste. Today, in the Western world, this can be practically applied by using toilets and a septic tank, or sewage treatment systems. Throughout history, mankind has not always heeded the advice of Moses and has suffered the consequences of severe diarrhea, dysentery, and other gastrointestinal maladies including severe dehydration, which could even lead to death. On a historic note, it is believed that S. dysenteriae, E. coli, and amoebic dysentery were responsible for nearly 2 million cases of diarrhea (Fig. 6) during the American Civil War. According to Hart (2004), there were 1,739,135 cases; including 44,558 deaths due to Bacillary Dysentery.
Theodor Escherich
Theodor Escherich (1857–1911) was a pediatrician and a professor at German and Austrian universities (Fig. 2). He discovered the bacterium Bacillus coli in children who suffered intestinal diseases. He was driven by a belief that studying bacteriology could solve many of the ailments suffered by children. He studied infant feces for the causative organism and amongst his inventory of microbes was a quick-growing rod-shaped bacterium he called initially Bacillus communis coli. It was renamed E. coli after him, in 1919.
The Coliform Kind Becomes Corrupt (A Type of Germ Genesis)
There are about 500 E. coli strains. Most strains are very good creations and considered to be mutualistic, symbionts (mutually helpful organisms) with humans and many other creatures. As we write this article, more than 43 genomes of E. coli have been sequenced. With every new strain, microbiologists discover new genes that are unique to each strain. Each strain shares some DNA with other strains (maybe 30% of 4–5 million base pairs) and Shigella. The pangenome of E. coli is about 11,000 genes; this is the total set of genes for all E. coli strains currently known (Zimmer 2008 p. 117).
Some E. coli cause traveler’s diarrhea and others cause even more severe dysentery. New strains continue to “pop up” with minor variations of the original kind. Most recently, E. coli O145 has been the cause of foodborne illness in those who ate Romaine lettuce. Pathogenic E. coli are usually divided into 5 categories (Nester et al. 2009):
- Enterotoxigenic E. coli (ETEC)
- Enteropathogenic E. coli (EPEC)
- Enteroinvasive E. coli (EIEC)
- Enteroaggregative E. coli (EAEC)
- Enterohemorrhagic E. coli (EHEC), also called Shiga toxin-producing (STEC)
Two strains (E. coli K-12 and O157:H7) are well known and represent the extreme variation. These two show how diverse and adaptive E. coli is as a created “kind.” K-12 is harmless under normal, healthy conditions. It is a “laboratory pet” and can easily die in competition with other lab fungi and bacteria. It might be considered a household “kitten” that has been “declawed.”
Fig. 7. E. coli O157:H7 (CDC image # 10068) Virulence factors include fimbraieae, adhesions, type III secretion system; and produce Shiga-like toxin.
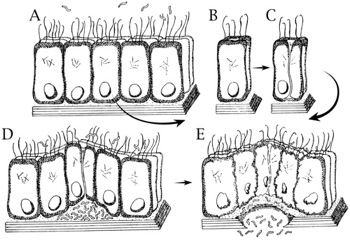
Fig. 8. Post-Fall consequence. Boundaries broken and bacterial invade. The establishment of infection frequently involves an invasion of host tissues when the boundaries are broken. In the lettered sequence, A through E, there is a progressive break in the basement membrane of epithelial tissue because of bacteria, like pathogenic E. coli strains that release enterotoxins. This poison may disrupt Peyer’s patch (a lymphoid tissue in the GI tract) and/or the intestinal wall barrier. This disruption makes room for more bacteria to colonize the tissue. Dysentery, typhoid fever, and bloody diarrhea are possible. In addition, fatal diseases like hemolytic uremic syndrome are possible if toxins from E. coli O157:H7 spread in the bloodstream from the GI tract to the kidney. Injury, disease, and even death are possible when boundaries like these are broken (Adapted from Gillen, Sherwin and Knowles 2001, p. 108).
On the other hand, E. coli O157:H7 is a “lion” of a bacterium (Figs. 7 and 8). It has the ability to inject toxins into our cells (specifically kidney and intestinal cells), cause internal bleeding, shut down vital organs, and kill people. This usually results in hemolytic uremic syndrome and a critical care situation in a hospital emergency room. This strain was first recognized in 1982, and since then has emerged as a public health problem. It is now one of the leading causes of diarrhea worldwide. In 1996, some 9,000 people in Japan became ill and seven died. The recent outbreaks of E. coli O157:H7 in the U.S., associated with contamination of spinach, undercooked meat and unpasteurized beverages (like apple juice), have alerted public health officials that new methods of testing for bacteria in food must be developed.
Each E. coli strain relies on a network of genes and proteins to survive in a particular niche. These probably have a recent common ancestor probably a 20th century ancestor! Something changed, and this is the so-called “stepwise evolution.” It is conceivable that after the Edenic Fall, some of the enteric bacterial kind became corrupted by the addition of genetic material (from horizontal transfer of genetic material from other bacteria); as well as from the loss of chromosomal DNA. Modification of genetic information in the Escherichia kind and displacement thus probably led to its pathogenicity (Fig. 5). (Displacement occurs when microbes that were originally designed to perform beneficial functions in a specific location, spread to other places after the Fall and began to cause disease.) Today, most Escherichia have only a trace of such modification, and therefore most remain beneficial. Conversely, a few strains of Escherichia show large genetic components that seem to have been added and a few genes that have been lost.
The pathway to pathogenicity for Escherichia appears to result from several major traits that have been added, including genes that code for a toxin, virulent enzymes, and traits that help bacteria avoid phagocytosis by white blood cells. These traits may have come from transfer of genes from neighboring bacteria.
Gram-Negative Outer Membrane
E. coli are Gram-negative bacteria (Fig. 1). Some of the same cell wall properties that facilitate their red color with stain also contribute to their variation and virulence. Gram-negative bacteria contain lipopolysaccharides in their cell walls that sometimes contain poisonous “endotoxins.” The cell walls of Gram-negative bacteria are more complex chemically than those of Gram-positive organisms. The two most critical antigens in E. coli that contribute to its pathogenicity and virulence are the O and H antigens. The O polysaccharide antigen is found within the cell membrane. The H antigen is part of the flagella. There are 50 plus strains of E. coli due to various H antigens. E. coli have fimbriae, or hair-like extensions, to help them attach to the intestine and eventually form a biofilm. Without this attachment and subsequent multiplication within a biofilm, there is no disease. In today’s post-fall world, E. coli O157:H7 bacteria are far more virulent than E. coli K-12.
Pathway to Pathogenicity
The written history of E. coli is complex (Fig. 4) and has been tracked by thousands of scientists who are “biological genealogists.” This history begins with Theodor Escherich in 1885. Today, hundreds of microbiologists follow his diligent and painstaking work. However, they now look primarily at DNA and the genetic “record” of generations of E. coli. When a mutation or horizontal transfer occurs in E. coli, it is frequently passed down to its offspring. That mutation or other change can sometimes serve as a genetic marker that tells microbiologists that particular bacteria may be related (Tortora, Funke, and Case 2010; Zimmer 2008).
E. coli has about 4000 genes and roughly 5 million DNA base pairs (BP). (In contrast, humans have 20,000 genes.) Two strains (E. coli K-12 and O157:H7) share about 75% (3/4) of the same genes (Table 2). It appears that E. coli O157:H7 has gained about 800,000 DNA base pairs, many of which are factors that led to its pathogenicity and its ability to avoid the body’s defense system (Seifert and DiRita 2006). As we examine many other important pathogenic bacteria, it appears that a “recipe for corruption” is: the addition of DNA from other bacteria, the loss of DNA, and the mixing of “foreign seed” (DNA from other microbes) with the original kind (“seed” DNA of the original type of E. coli). This change in DNA composition may be beneficial to the bacteria for a season.
Table 2. Comparison of the Two Major E. coli Types
| Strain | DNA Size (MBP) | Number of Genes | Comments |
|---|---|---|---|
| K-12 | 4.64 | 4,377 | Nonpathogen; the “laboratory pet” |
| O157:H7 | 5.44 | 5,416 | Pathogen; strain that is extremely virulent to humans; has 1,346 genes not found in E. coli K-12 |
Virulence seems to commonly arise via mechanisms of adding DNA through conjugation, transformation, and transduction, and loss of DNA encoding metabolic information (Figure 4). For example, bacteriophages (viruses that infect E. coli in a process known as transduction) are known to carry incomplete or corrupted genes that are inserted into the genome of the host they infect. In addition, pathogenicity islands (Purdom 2009) have apparently been transferred via conjugation from other bacteria, thus adding new virulence factors. The most significant lateral transfer appears to be the addition of chromosomal DNA cassettes (pathogenicity islands) from neighboring bacteria (Seifert and DiRita 2006).
The Hemolytic Uremic Syndrome and Kidney “Shutdown”
Hemolytic-uremic syndrome (HUS) often occurs after a gastrointestinal infection with enteric bacteria like E. coli O157:H7 (Figs. 7 and 8), S. dysenteriae or Salmonella. Hemolytic-uremic syndrome (HUS) is a disorder that usually occurs when an infection in the digestive system produces toxic substances that destroy red blood cells. It often affects the kidneys and is most common in young children. Although at one time it was very rare, it is increasing and is now the most common cause of acute kidney failure in children. Several large outbreaks in the 1990s and 2000s were linked to undercooked hamburger meat contaminated with E. coli, or with children drinking contaminated pool water. HUS often begins with vomiting and diarrhea, either of which may be bloody. Within a week, the person may become weak and irritable. Persons with this condition may urinate far less than normal; in fact, urine output may almost stop. Furthermore, red blood cell destruction can lead to symptoms of anemia. This deadly condition may cause some to ask, “What mechanism caused some of these very good designs to go bad? Did the Creator directly modify E. coli, or did they change over time via mechanisms that were corrupted as a result of the Fall?”
Modification and Displacement of Original Kinds
Before answering these questions directly, we need some perspective on microbes, in general. Creation scientists would suggest that pathogenesis is a secondary state in nature. At least three possible changes seemed to have occurred and perhaps a combination of all three took place. These include modification, displacement, and uncontrolled growth. The modification of the “original” genes (DNA that has been added, deleted, or mutated) is how a very good design may have been corrupted. This corrupted message is a result of the Curse. Dr. Francis (2009) defines these processes. Modification is one or more changes in the DNA that differs from the original kind. Microbes were physically modified—through genetic (DNA) addition, mixing, or deletion that lead to pathogenicity (Francis 2009). Displacement (Francis 2009) is where microbes were originally designed to perform beneficial functions in restricted anatomical or environmental locations, but after the Fall they spread to other places and began to cause disease. Some have uncontrolled growth—their numbers were meant (designed) to stay in safe ranges, but now they fluctuate and when overpopulated they cause disease or disruption of a once balanced system.
In earlier discussion within this article, changes have been assumed to have been based upon modification of bacterial genomes. Perhaps over time, the genome of the original E. coli (or ancestral prototype enteric bacterial type) changed, and this “good” bacteria, which provided benefit within its designed host was modified (addition or deletion of DNA) and then displaced. It was dispersed to an originally uninhabited host and the bacterial type was detrimental (disease-causing) in the new host. In the case of the E. coli that became strain O157:H7, the original strain may have originally been intended/created to live symbiotically in the gut of cattle, sheep, and other similar mammals. Recall that in a somewhat analogous way, man was not originally instructed to eat meat; humans were told to eat plants (fruits and vegetables; Genesis 1:29; Genesis 2:9). It was only after the worldwide Flood of Noah’s day, that the Creator told man that he could begin to eat meat (Genesis 9:1–4). After that, eating cattle, sheep, or similar mammals may have allowed the strains of E. coli from animals and humans to come into close contact. Movement of genes into the E. coli that inhabited the human gut from E. coli that inhabited the animal gut may have produced the O157:H7 strain that was pathogenic in humans.
Displacement of the Toxin Gene
E. coli O157:H7 normally lives in mammals other than humans. In contrast, most E. coli and Shigella strains are highly adapted to living in humans and only rarely in other animals. E. coli O157:H7 lives primarily in cattle, sheep and other farm animals. In humans, E. coli O157:H7 is deadly; but in farm animals it is harmless and sometimes even beneficial. E. coli O157:H7 may be designed to make “toxins” to help their animal hosts in some way. Microbiologists at the University of Idaho have found that sheep infected with E. coli O157:H7 do a better job of withstanding a cancer-causing virus than sheep without the strain. They hypothesize that in sheep, E. coli O157:H7 stimulate the animals’ immune system, or perhaps even triggers cells infected with a cancer-causing virus to undergo programmed (good) cell death before they form tumors. It is also possible that the bacterial toxins act as a defense for the bacteria themselves, i.e., when protozoans attack E. coli colonies, the E. coli O157:H7 strain fares better against predators. In cattle, E. coli O157:H7 also tends to colonize cattle more than E. coli K-12. This suggests that this strain although dangerous to humans may help cattle obtain enhanced benefits such as increased levels of vitamin K. In any case, E. coli O157:H7 does not appear to harm these farm animals.
So, is E. coli O157:H7 really new and evolving? Yes and No. The specific strain, E. coli O157:H7, may be really new (from about 1982); however, pathogenic strains that are like it are probably not new. E. coli has probably undergone significant change since its creation at the time of Genesis 1. God made all life (including E. coli) to adapt to changing conditions. Therefore, variation through time is the norm in microbes and all other creatures. Much as dogs, cats, roses, and people vary somewhat with environmental conditions, so do bacteria. In this fallen world, all creatures can cause problems for other creatures.
Summary and Conclusions
The mutualism among various organisms when viewed from the Christian worldview is evidence for creation, i.e., E. coli and human intestinal tissue are normally intimately in contact and extend the intestines’ interwoven complexity. E. coli produces Vitamin K, helps utilize vitamin B12, manufactures other nutrients, breaks down waste, and assists the immune system. It is part of a life-sustaining biomatrix within the intestine. It is yet another example of the organosubstrate theory of microbes and man. It is evidence of a loving Sustainer, Creator, and Maker of life. It is a biomatrix of two complex life forms that promote life for one another.
In my opinion whatever we may have to go through now is less than nothing compared with the magnificent future God has planned for us. The whole creation is on tiptoe to see the wonderful sight of the sons of God coming into their own. The world of creation cannot as yet see reality, not because it chooses to be blind, but because in God's purpose it has been so limited - yet it has been given hope. And the hope is that in the end the whole of created life will be rescued from the tyranny of change and decay, and have its share in that magnificent liberty which can only belong to the children of God! Romans 8:18–21, Phillips Translation.
In conclusion, the relationship between intestinal bacteria and humans is optimal for survival because of the ability for cooperation within this entire “package of parts.” In this case, it is a mutualistic, coordinated system of microbes working together with man’s gastrointestinal tract. It is adaptive in that it increases the survival of both humans and microbes. In fact, this remarkable unity between these magnificent microbes and our intestines can be logically derived as having its origin in a Creator. This remarkable cooperation is strong evidence for creation because it defies the laws of probability that two very different organisms can live together in such remarkable harmony. They extend the fearfully and wonderfully made design and interwoven complexity of humans (Gillen 2007). The dynamic digestive system with its interacting parts, the small intestine’s villi and the colon with their mutualistic bacteria, are part of the Creator’s wise design for the human body. No doubt these organisms were a part of the original blueprint for the human body. Since the Fall of man, however, there are a number of pathogens that invade the same niches normally occupied by harmless bacteria. Invading pathogens cause havoc in our body and, in extreme circumstances, may even cause death.
E. coli were made to adapt to a changing world. In order to “survive,” they have mutated, changed, and adapted (although they remain the same basic kind of bacteria). Sometimes in the process of changing, they cause disease in their host organism. They do not change to harm their hosts, but they sometimes do because they have themselves become subject to the Curse’s law of decay and corruption, as have their hosts.
No doubt, E. coli and other microbes will continue to adapt and change over time. This adaptation is and will always be “within their kind.” New diseases will come and go. However, the microbe itself has been designed for stability and continuity. E. coli are still a part of their original created kind. Someday, E. coli and man will be free from the Curse, and we believe that our new body will be free of germs. In the new heaven and earth, disease will be removed, and we will have no worry about pathogenic microbes (Revelation 22:2).
Since his Fall, man has struggled to conquer disease, one of the curses brought by sin. However only in the past two centuries has mankind made great strides curing and preventing diseases. Pasteur, Lister, Koch, Ross, and Reed worked from a creation perspective while trying to understand disease, to spare people from suffering from infections. Their lives were focused on experimental science as it related to infectious disease, but they were also influenced by a Judeo-Christian worldview. Many of the “founders” of modern-day microbiology held a worldview that was consistent with historic and traditional biblical teachings, including those concerning Creation, the Curse, Corruption, Contagion, and Christ (Gillen and Oliver 2009). If we help bring back a Biblical worldview, we may expect to again see even greater medical advances that may again increase mankind’s quality of life, and reduce suffering.
Until the Great Physician comes, be sure to wash your hands, clean your garden greens, and grill your hamburgers until they are well done! In this way, you honor the Creator through practical application of science and the Bible.
Acknowledgements
I would like to express my appreciation for the feedback of Georgia Purdom and Andrew J. Fabich on early drafts of the manuscript.
References
Bauman, R. 2009. Microbiology with diseases by body system, 2nd ed. San Francisco, CA: Pearson Benjamin/ Cummings Pub. Co.
Dininny. 2010. Regulators consider broadening testing for E. coli. Associated Press. www.signonsandiego.com/news/2010/jun/15/regulators-consider-broadening-testing-for-e-coli/ (Accessed June 28, 2010).
Escherich, T. 1885. Die Darmbakterien des Neugeboren und Sauglings Fortschr. Med. 3:515–522, 547–554.
Francis, J. W. 2003. The organosubstrate of life: A creationist perspective of microbes and viruses. In, Ivey, R. L. (ed.), Proceedings of the fifth international conference on creationism, pp.434–444. Pittsburgh: Creation Science Fellowship.
Francis, J. 2009. What about bacteria? In Ham, K. (editor). The New Answers Book 3. Green Forest, Arkansas: Master Books.
Gillen, A. L., Sherwin, F. and A. C. Knowles. 2001. The Human Body: An Intelligent Design, 2nd ed. St. Joseph, Missouri: Creation Research Society Books.
Gillen, A. L. 2006. Body by Design: Fearfully & Wonderfully Made. 5th printing. Green Forest, Arkansas: Master Books.
Gillen, A. L. 2007. The Genesis of Germs: Disease and the Coming Plagues in a Fallen World. Green Forest, Arkansas: Master Books.
Gillen, A. L. 2008. Microbes and the Days of Creation. Answers Research Journal 1 (2008):7-10.
Gillen, A. L. 2009. The Genesis of MRSA: A Modern Day “Leprosy” and Hospital Menace. Answers in Depth 4 no. 1.
Gillen, A. L. and J. D. Oliver. 2009. Creation and the Germ Theory: How a Biblical Worldview Helped Shape the View that Germs Make Us Sick. Answers in Depth 4 no. 1.
Hart, T. 2004. Microterrors: The Complete Guide to Bacterial, Viral and Fungal Infections That Threaten Our Health. Buffalo, New York: Firefly Books.
Nester, E. N., C. E. Roberts, N. N. Pearsall, D. G. Anderson, & M. T. Nester. 2009. Microbiology: A human perspective, 6th ed. Boston: WCB McGraw-Hill.
Purdom, G. 2009. The Role of Genomic Islands, Mutation, and Displacement in the Origin of Bacterial Pathogenicity. Answers Research Journal 2 (2009): 133–150.
Seifert, H. S. and V. J. DiRita. 2006. Evolution of microbial pathogens. Washington, DC: ASM Press.
Talaro, K. P. 2008. Foundations in microbiology, 6th ed. Boston: WCB McGraw-Hill.
Tortora, G. J., B. R. Funke, and C. L. Case. 2010. Microbiology, an introduction, 10th ed. San Francisco, California: Pearson Benjamin/Cummings Pub. Co.
Zimmer, C. 2008. E. coli and the New Science of Life. New York: Pantheon Books.
Glossary***
Adaptation package – biological organisms are more than the sum of individual structures; their ability to function successfully is due to an entire “package of parts.”
Biofilm – an aggregate of microbe cells that are “stuck” to each other and/or to a surface. These adherent cells are frequently embedded within a self-produced matrix of extracellular substance. Biofilm is also referred to as “slime,” an interwoven mix of DNA, proteins and polysaccharides. It is the “matrix” of life.
Coliform – group of gram-negative bacilli from the Enterobacteriaceae family that produces gas via lactose fermentation. Their presence is commonly considered an indication of fecal contamination. E. coli is an example of a coliform.
Conjugation – a temporary union of two organisms for the purpose of DNA transfer.
Dysentery- an inflammatory disorder of the intestine, especially of the colon, that results in severe diarrhea containing mucus and/or blood in the feces with fever and abdominal pain (simplified from Wikipedia).
E. coli, Escherichia coli – an enteric bacterial species used in many biological research projects which inhabits the intestines of humans and other animals. Proliferation can cause newborn meningitis, diarrhea, and urinary infections.
Enteric bacteria – bacteria that reside in the intestinal tract. Most of these bacteria are not harmful to the human body and many are mutualistic with humans, providing valuable vitamins and breaking down nutrients.
Escherichia – bacteria genus belonging to the family Enterobacteriaceae. E. coli is the most common isolate of this genus and is a part of the normal flora of the intestinal tract. It is, however, capable of causing illnesses ranging from mild food poisoning to severe kidney damage, and even death. The pathogenic serovars (types) of E. coli are divided into four groups, whose identification usually require serological methods.
Lateral (horizontal) gene transfer – any process in which an organism transfers DNA to another cell that is not its offspring.
Mutualism – a relationship between two or more organisms of different species where all benefit from the association.
Pangenome- the full set of genes for a bacterial type (i.e., E. coli) that is contained within all strains. Pathogenicity island – a cassette of genes involved causing infectious disease. This is not the “evolution” of new chromosomal DNA, but was an acquisition through lateral gene transfer.
Phagocytosis – the process by which a cell engulfs foreign substances or other cells; the particle containing vacuole fuses with a lysosome whose enzymes digest the food. An example of this is a macrophage eating a bacterium in the blood.
Shigella dysenteriae – a species of the rod-shaped bacterial genus Shigella. Shigella can cause bacillary dysentery. Shigellae are Gram-negative, facultatively anaerobic, non-motile bacteria. S. dysenteriae, spread by contaminated water and food, causes the most severe dysentery because of its potent and deadly Shiga toxin, but other species (like E. coli) may also be dysentery agents (simplified from Wikipedia).
Symbiosis – a close relationship between two or more species where they live together for a prolonged period of time.
Transduction – the process in which infection by a virus results in DNA being transferred from one bacterium to another.
Transformation – the transfer of a DNA segment from a nonfunctional donor cell to that of a functional recipient cell.
*** Unless stated, otherwise, terms are taken from Gillen (2007).

Footnotes
- The term, creation biologist, is used in the broader sense. Although there is strong historical evidence that each of these men had a creation worldview, their exact stance (young-earth, old-earth, progressive creation) is not known.
- Wove together – this implies a mutualistic relationship between two distinct organisms, not amosaic of one.
- Creation, Curse, Corruption, and Contagion: Adam’s sin ushered death, sickness, and sorrow into the once-perfect creation (Romans 5:12). God also pronounced a curse on the world, changing it completely (Genesis 3; Romans 8:20–22). As a result, the world that we now live in is merely a decaying remnant—a corruption—of the beautiful, righteous world that Adam and Eve originally called home. Some of the original very good microbes became pathogens (via modification or displacement), and the earth was filled with germs. These germs became responsible for contagious diseases (i.e. contagion) (Leviticus 13; Gillen 2007). For more information on Answers in Genesis’s 7 Cs, see https://answersingenesis.org/bible-history/so-what-are-the-7-cs-anyway/.

Support the creation/gospel message by donating or getting involved!

Answers in Genesis is an apologetics ministry, dedicated to helping Christians defend their faith and proclaim the good news of Jesus Christ.
- Customer Service 800.778.3390
- Available Monday–Friday | 9 AM–5 PM ET
- © 2026 Answers in Genesis