
The views expressed in this paper are those of the writer(s) and are not necessarily those of the ARJ Editor or Answers in Genesis.
Abstract
Young-earth creationists rightly consider that Neanderthals were human, but are divided on various issues, mainly because of the opposing views of Jack Cuozzo, author of Buried Alive! and Marvin Lubenow, author of Bones of Contention. These differences include Neanderthal lifespan, causes of the Neanderthal distinctive morphology, why the Neanderthals disappeared, length of Neanderthal children’s maturation time, whether we can place Neanderthals in biblical history, and the related issue of whether Homo erectus was human. In addition, Neanderthals are implicated in matters that include the Babel dispersion, earth’s breakup, DNA, patterns of world population, the Ice Age, archaeology, and historical timeline.

Keywords: Neanderthals, Homo neanderthalensis, Neanderthal morphology, Neanderthal DNA, Homo erectus, timeline, human lifespan, ancient DNA, Ice Age, ancient history, Babel dispersion.
Introduction
Homo neanderthalensis was the scientific name given to an unusual ancient fossil (later to be called Neanderthal Man) found in the Neander Valley near Dusseldorf, Germany, in 1856. It was later realized that fossils of H. neanderthalensis had been discovered earlier in Engis, Belgium, in 1830, and in Forbes’ Quarry, Gibraltar, in 1848. (For an extensive history of Neanderthal finds, see, for example, Trinkaus and Shipman 1993).
At that early time, these fossils were considered to be ancient, primitive humans by some (who called them “ape-man”), or diseased modern humans by others, but nonetheless human (Regal 2004, pp. 38–43). Reconstructions of what Neanderthals might have looked like when alive gave them a very satisfactory ape-like appearance (for example, fig. 1). In 1908, Neanderthal as a primitive, brutish, caveman was literally invented by Marcellin Boule of France (Regal 2004, pp. 51–52). That image of the Neanderthals was to persist for the next 50 years (Drell 2000; Schrenk and Muller 2008).
It has been generally conceded by evolutionists, however reluctantly, that they would have to accept that Neanderthals were as human as we were (Lewin 1999, pp. 156–163). But evolutionists haven’t given up entirely without a struggle, and they remain ambivalent about the Neanderthals. Hints of the evolutionist difficulty with considering Neanderthals entirely human keep surfacing, as in questions of whether they could really talk like us, for instance (Hoffecker 2005; Krause et al. 2007a; Swaminathan 2007). Speth (2004) found it necessary to chide his fellow scientists for convicting the Neanderthals of gross mental incompetence without adequate proof (“By most recent accounts, Neanderthals would have had considerable difficulty chewing gum and walking at the same time.”).

Fig. 1. First reconstruction of a Neanderthal man, by Schaaffhausen, 1888 (Neanderthal 2009).
Young-earth creationists, meanwhile, were not at all reluctant to recognize Neanderthals as human (Oard 2003a; Phillips 2000; Robertson and Sarfati 2003); after all, they had known from the beginning that there was no such thing as an ape-man. Lubenow pointed out that at several sites Neanderthals and modern humans were buried together, which he considered to be strong evidence that Neanderthals were of our species, because “In all of life, few desires are stronger than the desire to be buried with one’s own people” (Lubenow 2004, p. 254).
But there remained the serious job of examining this Neanderthal man. Who was he? On this question, creationists have been somewhat less than unified in their answers.
The Root of the Disagreement: Cuozzo versus Lubenow
Currently, the most widely accepted creationist view of the Neanderthals is that they were very early nomadic humans, probably one of the tribes that departed from Babel in the dispersion (for example, Oard 2003b, Robertson and Sarfati 2003). Their unique skull characteristics were possibly the result of family genetic traits and/or poor diet and lack of sunshine during the Ice Age, or perhaps disease. This is essentially the view of the Neanderthals that is described by Marvin Lubenow in his influential book, Bones of Contention (Lubenow 2004), as well as in various articles (for example, Lubenow 1998, 2000, 2006, 2007). Lubenow, theologian and anthropologist, had been studying fossils for many years and his conclusions seemed solid enough; creationist writers have been quite willing to subscribe to his ideas (for example, Oard 2009; Parker 2006; Purdom 2007).
But creationists were not totally unanimous in following the party line on Neanderthals. Maverick Jack Cuozzo, an orthodontist, had been studying Neanderthal skulls, and had authored some papers (for example, Cuozzo 1987, 1991, 1994). He then published his scientific research in a book on Neanderthals, Buried Alive! (Cuozzo 1998a); and presented a paper on computer projections of human skull changes with age at the International Conference on Creationism that same year (Cuozzo 1998b).
Cuozzo’s work represented a new research concept with regard to the scientific methods used in the study of ancient fossils. Whereas everybody else merely examined the Neanderthal skulls as they were at a point in time, Cuozzo looked at the skulls (using X-rays) as changing continuously from the moment of birth right on through to death. He had come to some radical conclusions: Neanderthals were ancient people who had developed their unique morphological characteristics (appearance) because they lived to an age of several hundred years, and, in addition, they had matured to adulthood very slowly. The significance of Cuozzo’s work did not appear to sink in among creationists, who have largely ignored his ideas.1 Creationists who have taken heed include Beasley (1992), Murdock (2004), and Robbins (2009).
Although most creationists currently accept the Lubenow view of Neanderthals, they admit that there are questions that remain. Where did the Neanderthals come from and where did they go? Why did they look as they did? Why did Neanderthals and modern humans appear to live side by side for long periods (or did they)? Why were Neanderthals buried with modern humans? Why did the robust Neanderthals disappear so suddenly? Do we carry any of their DNA or did their line go extinct? Why did young Neanderthals not have browridges? Where did the Neanderthals fit into biblical history? Were Neanderthals one of the groups dispersed from Babel? And where did Homo erectus fit into the picture?
Indeed, accepting H. erectus as human, as the majority of creationists do now, was a concept that raised its own set of questions. Where did H. erectus fit into biblical history? Where did H. erectus come from and where did he go? Do we carry H. erectus genes? Was there really a wide diversity of human genes immediately post Flood? If so, how did this diversity happen so soon after the human genetic bottleneck of the Ark? Should there not have been less diversity immediately after the Flood, with diversity increasing as time went on? How did H. erectus relate to the Neanderthals? Why are there no H. erectus burials (Homo erectus 2009b)?
Creationists as a whole did not seem to have answers for all these questions.
Homo erectus: Human or Not?
As we shall see later on, the status of H. erectus (whether human or not) has a direct bearing on some aspects of the Neanderthals. Therefore, at this point we will leave the Neanderthals temporarily to take a detour through the rather swampy territory of H. erectus.
It has been argued that H. erectus, as a class of fossils, was artificially manufactured by evolutionists to serve their purposes, and that H. erectus doesn’t really exist; therefore, as creationists, we should simply classify ancient fossils as either human or ape, and that should settle that (Bowden 1988). On the other hand, for a class of fossils that doesn’t exist, H. erectus has been getting mentioned quite regularly by creationists in their literature (for example, Gish 1985, pp. 180–203; Lubenow 2004; Mehlert 1994; Taylor 2008; Wieland 2002; Woodmorappe 2000.
Homo erectus began its rather checkered career as Pithecanthropus erectus (meaning “upright ape man”) with discovery of the first specimens in 1891 and 1892 in Java, Indonesia, by Eugene Dubois. In 1951, after a circuitous route of giving other names to various similar successive finds, this class of fossils was renamed Homo erectus (Lewin 1999, pp. 138–144).
Early creationists were generous about admitting H. erectus to the human dinner table. Rusch (1964), for instance, believed that not only H. erectus but even earlier forms of fossils in the claimed evolutionary chain were human. Tinkle (1968) believed that all apes had canine teeth (a shaky assumption), and that any fossil that did not have canine teeth had to be human; he therefore included as human some rather doubtful candidates. Shaw (1970) considered all homo-type fossils human, except possibly the Australopithecines, of which he was uncertain.
Then came a change in creationist thinking. According to Mehlert (1994),
The Javan and Peking forms of H. erectus in particular came under considerable attack by creationists in the 1970s and 1980s. The thrust of these critiques was that all H. erectus forms were extremely ape-like and even possibly fraudulent.
(See also Bowden 1981, pp. 78–148; Gish 1985; Johnson 1982, pp. 40–45.)
But the pendulum was to swing again. This was partly caused by discovery of the skeleton called Turkana Boy (KHM-WT-15000) in Africa in 1984 (Lewin 1999, p. 120), causing creationists to declare that now they would simply have to believe that H. erectus was human (Mehlert 1994). In his revised book, Lubenow (2004) pushed very hard on this theme, devoting almost 80 pages to defending H. erectus as fully human, including the controversial Javan and Peking fossils. Other creationists have willingly followed suit (for example, Line 2005; Wieland 2002; Wise 2005; Wood 2008; Woodmorappe 2000).
Even the tiny, meter-high Homo floresiensis, nicknamed the “hobbit,” discovered in Flores, Indonesia, in 2003 (Brown et al. 2004), was hailed as human by creationists (Wieland 2008), although evolutionists were wary (Argue et al. 2009; Baab and McNulty 2008; Larson et al. 2007). Just over three feet high, with a head the size of a grapefruit, this so-called human presented some puzzles, especially how it got so small. This prompted theories about such factors as disease, insular dwarfing, elevated climate temperature and CO2, and iodine deficiency (Tyler 2005).
The subject of human cranial capacity now needed to be addressed. Although creationists accepted that the average capacity of the H. erectus skulls was considerably smaller than that of modern humans (Homo erectus 2009a), this really small Flores skull did rather push the limits. Happily, Jue (1990) had earlier reassured creationists that “what is now known of modern man, there is no relationship between cranial capacity and intelligence.” In other words, merely because a fossil had a very small brain, this did not preclude it from being human. Everyone could relax.2
While most creationists were busily turning all H. erectus into members of the human race, there was another voice trying to make itself heard. Jack Cuozzo did not believe that H. erectus was human. Homo erectus, he said, was an ape—an ancient, advanced, ape, now extinct. “Apes probably were more complex at an early time in earth history, had more abilities, and might have been able to walk close to upright . . .” (Cuozzo 1998, p. 101). This tied in with Cuozzo’s strong belief, reiterated throughout his book, that all creation had originally been greatly superior to what it is now; and because of degeneration that set in after the fall in the Garden of Eden, all creatures had deteriorated over the millennia since then, or had gone extinct. In addition, he wrote, “. . . a detailed study of H. erectus shows very clearly it was not human, starting with the fact that there is no record of them ever burying their dead.”3 He went on,
I have an accurate cast from Africa of Nariokotome4 Boy’s skull and have done a diagnostic orthodontic set-up of its occlusion (upper and lower teeth in occlusion) and it is very ape-like . . and have heard its discoverer Alan Walker5 . . . enumerate the nonhuman features.
Also, with respect to the so-called hobbit, “My impression of it (the H. floresiensis fossil) is a complex small ape.” (Cuozzo 2009, private communications). It was clear that Cuozzo was not going to go along with the rest of the creationists on the matter of the various fossils that made up the class of H. erectus. Meanwhile, the changing creationist opinions on the status of H. erectus fossils have not gone unnoticed in the evolutionist camp. Foley (2008) illustrates the problem in a table of six H. erectus skulls and the varying creationist opinions on them.6 As Foley says, “although creationists are adamant that none of these are transitional and all are either apes or humans,7 they are not able to agree on which are which” (italics are Foley’s). To punch his point, he notes creationists who have flip-flopped, and also three examples of erectus fossils on which creationists can’t make up their minds. On the whole, this entire matter of the status of H. erectus has been rather embarrassing to creationists; if we are honest, we must acknowledge that there has been a problem in proving that we can tell the difference between fossil humans and nonhumans.
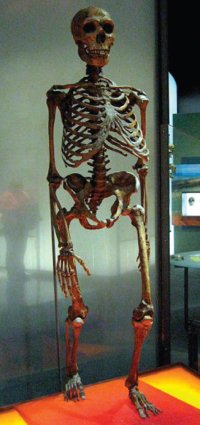
Fig. 2. Neanderthal skeleton on display at the American Museum of Natural History in New York. Photo by Claire Houck (Neanderthal 2009).
Why Are Creationists (and Evolutionists) Having Difficulty Distinguishing Between Early Apes and Humans?
It might seem that it would be obvious whether or not a skull or skeleton is that of a human like us. After all, live humans today are quite easily distinguished from all forms of apes. But without flesh and hair, the skeletal remains are more difficult to identify, as is clear from the foregoing discussion. For a comparison of these two taxons (groups), see Figs. 2, 3, and 4 (H. neanderthalensis) and Figs. 5 and 6 (H. erectus). The apparent similarities between H. neanderthalensis and H. erectus are striking.
One feature that is a source of confusion for both the evolutionists and creationists is bipedality (ability to walk on two feet). Indeed, Murdock (2006) blames evolutionists like Richard Leakey for using bipedality to confuse the issue of what constitutes a human. While most of Leakey’s evolutionist colleagues were using “hominid” to refer to all ancestral human species, Leakey unabashedly stretched the definition of “human” to all “apes that walked upright—bipedal apes.” And not only H. erectus were bipedal, he said, but also earlier species in the evolutionary line that led to H. sapiens (Leakey 1996, pp. xiii, xiv). On the creationist side, some have believed that only humans could have walked upright on two feet (for example, Wieland 1994), and therefore any being (such as H. erectus) that walked this way must have been human. However, there is strong support for bipedal, upright apes in ancient times, not only from Cuozzo (1998, p. 101), as noted earlier, but also from creationist paleoanthropologist Murdock (2006, p. 108) who does not mince words: “There has been unwillingness by some to accept the possibility that some apes walked upright in the past.” Murdock goes on to say,

Fig. 3. La Chapelle aux Saints (Neanderthal) skull, discovered in France in 1908 (Neanderthal 2009).
These apes walked upright, not because we share a common ancestor, but because we share a common designer. They must also be viewed as more complex versions of extant apes (having not suffered as many years of the curse) (Murdock 2006, p. 108).
Clearly, bipedality did not necessarily make H. erectus human (using the creationist definition of “human”).
Another noticeable source of confusion about the status of H. erectus is the prominent browridges of both H. erectus and Neanderthal; these browridges do admittedly make the skulls look similar, as shown in figs 4 and 6. However, we need to be cautious about this, because there are many fossils that are definitely not human that also have browridges; browridges alone do not determine humanness. This is well illustrated by an online photo8 of seven skulls in a row: only one (H. sapiens) has no browridges. All the others do, including a Neanderthal, a member of H. erectus, two Australopithecus, a gorilla, and a chimpanzee. One ape missing from this lineup is the orangutan (Pongo); like humans, these lack prominent browridges (Brow ridge n.d.).
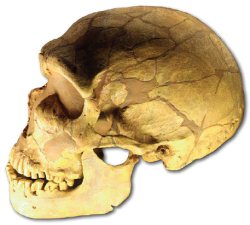
Fig. 4. La Ferrassie (Neanderthal) skull, discovered in France 1909 (Neanderthal 2009).
Custance (1968) began a long paper on human fossils by castigating evolutionist Howell (1967, p. 85) for writing that “Man is a primate and within the order of Primates is most closely related to the living African anthropoid apes.” “It is not at all safe to assume that all ‘look-alikes’ are related,” said Custance, calling Howell “purely presumptive” in stating this as a fact without proof. Forty years later, the two sides of this question remain about the same.
Those who consider H. erectus to be a Neanderthal look-alike would do well to look to the science of tooth development studies. Evolutionists Dean et al. (2001) have found that H. erectus and various other homo species had quite a different kind of tooth development than modern humans and Neanderthals; they consider this significant, stating that “Brain size, age at first reproduction, lifespan and other life-history traits correlate tightly with dental development.” This supports Cuozzo’s view that H. erectus and Neanderthal fall into different categories.

Fig. 5. Turkana Boy (H. erectus) skeleton on display at the American Museum of Natural History in New York. Photo by Claire Houck (Turkana Boy 2009).
Implications of Status of Homo erectus for Neanderthals
Creationists are offered a choice between two opposing views on H. erectus, whether human or not. They might well ask whether it really matters one way or the other.
What we need to understand is that the status of H. erectus has some important implications for Neanderthals. This starts with the number of different groups of very early people that we know about. If we believe that H. erectus was human, we have two possible candidates for peoples that could have left Babel in the dispersion of Genesis 11, Neanderthals and H. erectus. But Genesis 10 would seem to indicate that there were more groups than this. If so, who were they?
Consideration of this question logically leads to the conclusions of Wise, who says that since there must have been more peoples than this leaving Babel, there must be whole groups of early people who have disappeared, whether in natural catastrophe, warfare, or whatever (Wise 2005). Indeed, Wise goes as far as saying that H. erectus may have been the only humans at Babel; in his proposed scenario, Neanderthals would have possibly developed later. This raises the question whether this accords with what the Bible says in Genesis 10, where it lists a lot of different peoples, but nothing about disasters or eradicating any of them.
Also, as Wood points out, if H. erectus were human, this would also indicate that there was a wider range of physical appearance, and therefore genetic diversity, immediately after the Flood than we have today (Wood 2008). But in the view of others, there should have been less human diversity immediately after the two genetic bottlenecks of the Flood and the Babel dispersion, with diversity increasing as time went on (Ashcraft 2004; Nelson 2004).
Those problems are all solved if, like Cuozzo (1998, p. 101), we put H. erectus into the category of apes. This suggests that all the people who left Babel were Neanderthals who spread out in many directions to form the nations of Genesis 10; moreover, all peoples now living on earth are descended from Neanderthals. This theme will be developed in the next section on Neanderthal lifespan.

Fig. 6. Homo erectus skull on display at the Museum of Natural History, Ann Arbor, Michigan. Photo by Thomas Roche (Homo erectus 2009b).
If all the people who dispersed from Babel were Neanderthals, the various groups would have developed genetic changes as time went on, eventually to produce all the peoples that we see today. Recently, secular scientists have started to suggest exactly this, that the Neanderthals were not a homogeneous population, but that they diverged genetically in various geographical areas of the world where they lived (Fabre, Condemi, and Degioanni 2009). This would be a logical outcome of a worldwide Neanderthal dispersion. However, the reliability of this kind of testing of ancient DNA is questioned by some, a matter that will be discussed later in this paper.
Implications of Long Neanderthal Lifespan
As noted earlier, Cuozzo showed by his X-ray studies of Neanderthal skulls that these were people who must have lived for hundreds of years (Cuozzo 1998a). Not only that, his computerized modeling of skull changes with age predicted development of exactly the kind of characteristics that these Neanderthal skulls displayed (Cuozzo 1998b). This meant that there had to have been some very, very old people somewhere in our history.
Of course, we don’t have to look very hard to find extraordinarily long-lived people, because they are in plain view in the early historical Genesis accounts. Between Creation and the Flood, there were people like Adam who lived 930 years and Methuselah who lived 969 years, to pick two examples out of the genealogy of Genesis 5. Noah lived 950 years in all, 600 before the Flood and 350 after it (Genesis 7:11, 9:28, 29). Lifespans were still quite long for some time after the Flood; generations of people after Noah lived for hundreds of years (Genesis 11:10–22). If we accept the Bible literally, we have to believe that these people actually lived that long.
From this we can deduce that, according to Cuozzo’s studies, all the ancient long-lived people of early Genesis who lived for hundreds of years could be classified as Neanderthals, including everyone from Adam through to the Flood and for some generations after the Flood. Exactly when people no longer lived long enough to develop Neanderthal characteristics, and what generation after the Flood would have been the last of what we would call Neanderthals, Cuozzo does not specify at this stage of his work. (For a discussion of what factors are involved in estimating the probable ages of Neanderthal skulls, see Cuozzo 1998, pp. 201–216).
By contrast, the current creationist view of Neanderthals as simply an ancient tribe of nomadic people, now extinct, does not offer any answers as to who the Neanderthals really were. Lubenow, in discussing Neanderthal burial practices, says,
. . . I do not wish to imply that Abraham or his ancestors . . . were Neandertals. What the relationship was—if any—between the people of Genesis and the Neandertals we do not know (Lubenow 2007).
Cuozzo would beg to disagree with that.
What Caused the Neanderthals’ Unique Morphological Characteristics?
While everyone generally recognized that the Neanderthals were human, it was clear that they weren’t exactly like us in appearance. As Oard says (1996, p. 58), “Neanderthal Man did have a peculiar look about him.” How Neanderthals came to have their unique physical characteristics has been a subject of much debate by evolutionists and creationists alike, with no consensus reached on either side.
Early on, Custance (1968), a creationist anthropologist, had suggested that “extreme old age would often tend to modify the skull towards the conventional man-ape form.” But on this he was ahead of his time.
Since the Neanderthals had lived during the Ice Age, scientists on both sides have assumed that climate could have been a contributing factor in their facial and other characteristics (Mellars 1996, p. 3; Oard 1996, p. 59). But rats raised in the cold in controlled laboratory experiments showed some craniofacial changes opposite to those that Neanderthals had; apparently it would be necessary to reconsider the climate theory (Rae et al. 2006). There were, in any case, other possibilities that presented themselves. An article by a geographer (Dobson 1998) had caused rather a storm of controversy when he suggested that Neanderthals were deformed because of iodine deficiencies in their diet; in spite of attempts to refute this idea by the well-known Trinkaus (Gugliotta 1999), Dobson did not back down. Another group applied statistical tests to show that it was genetic drift that explained cranial differences between Neanderthals and modern humans (Weaver, Roseman, and Stringer 2007).
Disease was considered a likely candidate by many. Bowden (1981) had earlier taken the view that European Neanderthals were degenerate humans who suffered from rickets and syphilis; his belief was influenced by the opinions of a 19th-century pathologist, Virchow, and Ivanhoe (1970). The influential Virchow, who died in 1902,
believed that Neanderthal man was a modern Homo sapiens, whose deformations were caused by rickets in childhood and arthritis later in life, with the flattened skull due to powerful blows to the head (Schultz 2008).
Ivanhoe (1970) thought that Neanderthals may have suffered from a vitamin D deficiency and that Virchow was right after all. Jaroncyk (2007) thought a whole group of factors could be involved:
Some of their characteristic skeletal features could therefore be attributed to their harsh life in a cold post-Flood climate, as well as to arthritis, rickets and genetic isolation.
Oard (2006a, p. 129) favored rickets and arthritis because of a lack of vitamin D from cloudy Ice-Age weather, plus genetic inbreeding. Creationist Acton (1978) believed that rickets, Paget’s disease, and syphilis were the most likely candidates for causing the Neanderthal characteristics. Conrad (1986), an evolutionist, took Acton up on this, essentially saying that even though Acton had many medical titles, and knew a great deal about his own specialty, he clearly did not know enough about Neanderthals. Lubenow (2004, p. 84) has been satisfied to accept that disease, especially rickets and syphilis, could be contributors to the Neanderthal morphology, along with other factors.
On the subject of Neanderthals and disease, Lubenow and Cuozzo are definitely not in agreement. Cuozzo refutes the idea of diseases in some detail (Cuozzo 1998, pp. 275–279), concluding,
I sincerely hope that these outlined diagnostic signs for rheumatoid arthritis, rickets, and congenital syphilis will finally put to rest the speculation of all those who have tried to explain away the Neanderthal features using these diseases.
Although many of the authors above believed that the Neanderthals’ distinctive browridges could be largely accounted for by disease, various scientists thought that the browridges were actually produced by forces of chewing that affected Neanderthal facial structure (for example, Spencer and Demes 1993). But others concluded that what they called “masticatory biomechanical adaptation” (chewing-caused changes) did not underlie the Neanderthal facial morphology (structure), and that alternative explanations should be pursued (O’Connor, Franciscus, and Holton 2004), whatever those alternatives were. Lubenow (2004, p. 83) cites Klein (1989, pp. 281–282), whose “plausible nonevolutionary explanation for most of the unique features of Neandertal morphology” was that they gripped things with their teeth, using the latter as a tool.
Cuozzo, however, concluded that the Neanderthal browridges had to have formed merely from old age and normal chewing. He arrived at this from his studies of Eskimo skulls from Koniak Island at the Smithsonian in the 1980s; those jaws showed lumps of bone inside, formed because of the Eskimos’ tough premodern diet. But the Neanderthals did not have these lumps of bone, nor the thick gonial (jaw) angle of those Eskimos, and therefore did not seem to have had an unusually tough diet (Cuozzo 1998, pp. 182–184, 229–230).
If browridges were the result of old age, it naturally follows that Neanderthal children did not have them. This is indeed the case, as is shown in Fig. 7. In early times, however, it was mistakenly believed that Neanderthal children did have facial features just like those of adults. On this Cuozzo (1998, p. 272) says, “Many human paleontologists thought it was natural for Neanderthal children to have miniature adult faces.” He cites Howell (1957).

Fig. 7. Reconstruction of a Neanderthal child. Anthropological Institute, University of Zurich (Neanderthal 2009).
Although creationists might think that the subject of facial bone changes with age is new, the medical scientific community at large has been quite aware of this for a long time; surveys of the literature on the subject go back as far as the 1860s (Albert, Ricanek, and Patterson 2007; Behrents 1985; Guagliardo 1982). Israel (1977), for instance, pointed out that as people age, different parts of the craniofacial skeleton will grow at different rates. It may be surprising how wide the subject is of interest. A paper on craniofacial changes with age aimed at plastic surgeons suggested that surgeons may want to alter underlying bone structure as well as skin on aging patients (Bartlett, Grossman, and Whitaker 1992). At an FBI (Federal Bureau of Investigation) conference, Milner, Neave, and Wilkinson (2001) made the point that “bone remains a highly dynamic tissue with active remodeling occurring through old age.” Albert, Ricanek, and Patterson (2007) note that their research was funded by the United States Department of Defense which was interested in “studies of automated face recognition, computer 3D modeling of faces, and computer adult age progression techniques.”
Unlike creationists in general, who have been slow to pick up on Cuozzo’s work, those in dental fields of expertise have understood it quite well; for example, Robbins (2009), a retired dentist, has published an article in Bible and Spade in which he explains the principles behind Cuozzo’s calculations of lifespans of long-lived people.
Why Did the Neanderthals Disappear?
The biggest puzzle, that neither the majority of creationists nor the evolutionists have been able to solve, is why the Neanderthals disappeared. The robust Neanderthals appeared to have everything going for them, all agree, and there is no visible reason why they should not have survived (Trinkaus 1978). Nonetheless, disappear they did, rather suddenly, before the end of the post-Flood Ice Age or, as evolutionists call it, the end of the last ice age9 (Van Andel and Davies 2004).
Everyone agrees that modern humans showed up on the world scene at approximately the same time that the Neanderthals disappeared; whether or not this timing was a coincidence is debated. Some evolutionists allow thousands of years for the two groups to overlap—how many thousand is a matter of intense discussion—because they have a lot of time at their disposal, and a few thousand years here or there are a mere trifle (see, for example, Lewin 1999, pp. 157, 165–166). Creationists obviously have far less historical time available to account for the Neanderthal disappearance and subsequent appearance of modern man; therefore they have to explain how this mysterious event could have happened so quickly. But the problem for both sides is the same: why did it happen?
The proposed explanations forwarded by evolutionists on the Neanderthal demise have been both varied and creative, and only a sampling of the rather large literature on this subject can be touched on here. The Neanderthals’ supposed inability to cope with climate change has been especially popular (Jimenez-Espejo et al. 2007); although the Neanderthals had been able to live through the Ice Age successfully, they apparently could not cope with the ending of this cold period. Also much discussed are losing out to modern humans in various kinds of competition (Banks et al. 2008; Hoffecker 2002; Shea 2001), intermarrying with moderns (Zilhão 2006), or possibly both (Miller 2001). But there are others. Carnieri (2006) suggests that anatomically modern humans in Europe ate a lot of seafood; this more healthful diet helped them outlive the largely carnivorous Neanderthals. Sorensen (n.d.) suggests that Homo sapiens, migrating out of Africa, brought infectious disease that killed off the Neanderthals. Kuhn and Stiner (2006) argue that because Neanderthals did not divide their labor between the sexes the way modern humans did, this gave the latter a survival advantage. A mathematician, using what he calls a “simple mathematical homogeneous model of competition,” has determined that extinction of the Neanderthals was unavoidable (Flores 1998). Economists have gotten into the act with a theory that the Neanderthals came out second best because modern humans were better at trade (Horan, Bulte, and Shogren 2005). A rather grisly version surfaced in reports that, finally, there was good evidence that the Neanderthals actually did practice cannibalism, as had been suspected (Sanders 1999); presumably we were to believe that, like the gingham dog and the calico cat (Field 1894), the Neanderthals simply ate each other up. Then a different angle on the alleged cannibalism was proposed: eating each other, especially the brains, might have caused spreading of a mad-cow-related disease that could have played a large part in wiping the Neanderthals out (Underdown 2008). More recently, news articles (for example, McKie 2009) trumpeted to the world that it was actually cannibalistic modern humans who ate the Neanderthals up; this was based on an interview with scientist Fernando Rozzi, head of a research team that had just published a paper (Rozzi et al. 2009) that cast doubt on what their leader was telling the press(!). According to a group of geneticists, the small population size of Neanderthals may have made them more vulnerable to extinction, whatever the causes (Briggs et al. 2009).
This is not an exhaustive list of the many possibilities that have been proposed. As one insightful science newswriter says, “Figuring out why Neanderthals died out and what they were like when alive have kept plenty of scientists busy” (ANI 2009). Mark Twain would have been quite impressed by how little hard evidence supports some of these papers. He wrote, “There is something fascinating about science. One gets such wholesale returns of conjecture out of such a trifling investment of fact” (Twain 1883). One might think he was talking about evolutionists’ papers on the disappearance of the Neanderthals.
Meanwhile, creationists’ explanations of the Neanderthal demise have seemed rather tame and tentative by comparison, nor has there been a noticeable rush to embrace many of the various theories offered by evolutionists. Even Lubenow, who is very definite about many other ideas in his book, glosses lightly over the matter of why creationists think the Neanderthals disappeared from view; indeed, he speculates that Neanderthals could have survived into fairly recent times (Lubenow 2004, p. 82). The creationist stance is exemplified by a recent online piece about an apparent Neanderthal stabbing (Human stabbed a Neanderthal, evidence suggests, 2009), that ended with the words,
The more interesting debate is whether Neanderthals went entirely extinct . . . or whether their genes survive in many modern Europeans, as some studies have suggested.
On the one side, Oard (2006a, p. 129) states that the Neanderthals “very likely” intermarried with Cro-Magnon man, who seemed to follow the Neanderthals into Europe some time later; and Sarfati (2004, p. 317) concludes that “. . . modern humans and Neandertals likely amalgamated in Europe.” But, on the other side, Wise (2008) claims that DNA evidence shows that we do not carry Neanderthal genes today; therefore Neanderthals went extinct without intermarrying with modern humans. He speculates that this extinction event could have occurred because of challenges of survival in the post-Flood earth, or from various kinds of human violence.
The creationist debate as to whether or not the Neanderthals mixed their genes with those of modern humans through marriage is mirrored by evolutionists (who prefer to talk about “interbreeding” or “admixing” or “cohabiting”). Their positions are entrenched on both sides of this fence. “It is becoming increasingly clear that the Neanderthals and their modern human successors did not mix and that the Neanderthals are an extinct side branch of humanity” (Klein 2003); see also Currat and Excoffier (2004) and Tattersall (2007). But on the opposite side of the question are Wolpoff et al. (2004), who specifically refute Klein (2003); and Trinkhaus (2007), who believes that paleoanthropology shows definitively that the Neanderthals and moderns interbred, and the case is closed. Not so, says Paabo (Morgan 2009), whose belief in DNA and genome mapping (Green et al. 2008) bring him down on the side of almost total lack of interbreeding between Neanderthals and later humans. There would appear to be practically no middle ground between the two camps.
Predictably, progressive (old-earth) creationist Hugh Ross much prefers the DNA “proof” that Neanderthals and modern humans did not interbreed10; this is an extremely important matter to Ross, because if it can be shown that they did intermix, this would be “fatal to the current Progressive Creationist model,” according to Line (2007).
Obviously creationists and evolutionists are grappling with the same questions. Lubenow’s remark that the disappearance of the Neanderthals is like the disappearance of the Cheshire cat (Carroll 1865), whose grin remains to taunt evolutionists (Lubenow 2004, p. 81), applies equally to creationists. Clearly, the matter of what caused the disappearance of the Neanderthals has not been clear at all.
A major problem with most of the proffered hypotheses on the Neanderthal extinction is the widening geographical distribution of Neanderthal sites that have been located in the past few years, a subject that will be discussed later in this paper. Many authors address extinction of the Neanderthals in Europe, for example, and then rather ignore the ones in more far-flung places. Did other Neanderthals in other places become extinct for the same reasons? The whole subject becomes more complicated as the very large distances involved make it increasingly difficult to assume that everything can be explained by merely saying that the Neanderthals were nomadic.
However, the problem of the demise of the Neanderthals goes away entirely if we accept that Cuozzo is correct in his conclusions that the Neanderthals were the post-Flood long-lived people who spread out from Babel in all directions. Their “disappearance” would have occurred when they no longer lived long enough to develop the distinctive Neanderthal characteristics.11 The modern humans who supposedly “replaced” the Neanderthals would be the descendants of the latter, who did not live as long as their ancestors. This not only makes the matter of the Neanderthal disappearance very simple and straightforward, it also explains why it happens that modern humans arose at around the same time as the Neanderthals disappeared; furthermore, this would be true in all parts of the world. Proponents of Occam’s razor (Occam’s razor 2009), often stated as “The simplest explanation is usually the best,” would recognize the Cuozzo explanation of the Neanderthal demise as a good one.
According to Cuozzo, we would expect that, with people’s decreasing lifespans as time went on, the Neanderthal characteristics would gradually lessen from generation to generation, and then disappear entirely. In fact, this is what we see in various archaeological discoveries, although these are usually interpreted as humans that are the result of intermarriage between the Neanderthal and modern peoples (except for the DNA proponents, who do not agree, and who propose other ideas). For example, excavations in Israel are claimed to show “continuous biological evolution from Neanderthal to anatomically modern Homo sapiens” (Jelinek 1982). Also, at the Neanderthal site in Romania, the human remains display a “mosaic of modern human and archaic and/or Neandertal features” according to the paper published on the find (Soficaru, Dobos, and Trinkaus 2006). Creationists have hailed this as exciting news and further evidence that the Neanderthals were fully human beings (Anderson 2006; Jaroncyk 2007).
It follows logically that Cuozzo’s work knocks out the underpinnings of the Ross old-earth belief system, since the Ross view of Neanderthals as animals without spirits is nullified.
Neanderthal DNA: What Does It Tell Us?
As we have seen in the previous section, genetics and paleoanthropology are pitted against each other in the matter of the Neanderthal disappearance. On the one side, the geneticists, leaning on mapping of the Neanderthal genome and DNA sequencing, do not believe that we carry any Neanderthal genes today, lending their support to extinction of the Neanderthals as a side group not in our direct lineage. On the other side, the paleoanthropologists, pointing to the fossils they have found, believe that there was mixing between the Neanderthals and our modern ancestors, and that we surely must have Neanderthal genes in our makeup, no matter what anyone says. If Cuozzo is right that we are all descended from Neanderthals, then the paleoanthropologists must be correct as well. But what of the DNA?
Recent publications by two creationist scientists may dampen enthusiasm for drawing firm conclusions from studies of Neanderthal DNA. Carter (2009), a geneticist, cautions that we need to be skeptical about evolutionary assumptions, accuracy of the mtDNA (mitochondrial DNA) sequence, and other factors that might lead to wrong conclusions. Criswell (2009), a molecular biologist, points out that, although DNA has been extracted from Neanderthals, this does not necessarily mean that the DNA is of very good quality, especially because of contamination of the samples and (not surprisingly) decay over the millennia. It is beyond the scope of this paper to cover this subject in detail; those interested in pursuing the matter of Neanderthal DNA in more depth are encouraged to read both of these papers.
DNA is one area where Cuozzo and Lubenow agree. The latter’s Chapter 23, “Technical Section: mtDNA Neandertal Park—A Catch 22,” is an excellent overview of the matter of DNA, including the political implications of the struggle that he calls “the molecules” versus “the fossils” (this chapter is not very technical in spite of its title) (Lubenow 2004). The bottom line is that Lubenow does not consider DNA to be a reliable source of scientific information. Cuozzo does not put any weight on the mtDNA results, either, stating that there are changes in mtDNA over a person’s lifetime, so that the old Neanderthals would have different mtDNA than the young ones; the genome of the ancient people would have been far less devolved than ours; and genetic mutations in mtDNA have been occurring much faster than previously believed (Cuozzo 2009). This theme of continuous human physical degeneration from the earliest people to the present has been ably developed by Sanford (2008), who attributes the shortening of our lifespan throughout the millennia to accumulation of damaging genetic mutations.
One more thing should be mentioned here. If these Neanderthals were extremely old when they died, it would not be surprising if they showed some old-age characteristics, such as arthritis, and some signs of trauma such as healed broken bones (Trinkaus 1978). Indeed, it is surprising that the skeletons of these old people were in such good shape overall at the time of their death. Writers go on and on about how strong these people were (for example, Trinkaus 1978). By comparison, we can see how much we humans have degenerated in the thousands of years since then; our own old people, who do not live nearly as long, show many signs of disease and deterioration in their old age. Degeneration of our genome has taken its very visible toll.
Slow Maturation of Neanderthal Children and Its timeline Implications
The fossils of young Neanderthals examined by Cuozzo displayed characteristics that he believed could only be explained by a very slow rate of maturation, compared to the rate of maturation of children today (Cuozzo 1998, 2003). Since modern humans are considered to be slow to mature in the general scheme of living beings, this meant that the Neanderthals’ rate would have been very slow indeed. According to Krogman (1972, p. 2),
Man has absolutely the most protracted period of infancy, childhood and juvenility of all forms of life. . . .Nearly thirty percent of his entire life-span is devoted to growing.
That percent would be considered a bit high today in countries where life expectancy at birth is now over 80 years, but many countries still have a far lower life expectancy (Life expectancy at birth 2008), thus increasing the percentage of total lifetime spent growing up.
But by contrast, most secular scientists believed that Neanderthals had matured exceptionally fast; the usual explanation was that chimpanzees (touted widely as “our closest living relatives”; see, for example, Lovgren 2005) also matured rapidly (Moskowitz 2008). Indeed, Rozzi and de Castro (2004) studied Neanderthal teeth and reached the conclusion that Neanderthals became adults by a mere 15 years old. Whether or not creationists as a whole have accepted an unusually short lifespan of Neanderthals (perhaps because of belief that climate and disease factors caused the Neanderthal morphology), and a short maturation time, is hard to say, as it is difficult to find much in print on this point. However, on the matter of exceptionally long-lived, slow-maturing Neanderthals, Cuozzo was swimming nearly alone against the current.12 That is, until a group of evolutionists (Ponce de Leon et al. 2008) concluded from a study of skeletons that Neanderthals probably matured more slowly (after infancy) than modern humans, and lived to a relatively ripe old age, considering the popular view of short-lived Neanderthals; thus giving Cuozzo at least some support from an unexpected quarter.
If Neanderthal children matured very slowly compared to us today, it follows logically that it took longer than now to reach adulthood. Since we are postulating that the Neanderthals were the old people of the Bible, we will start by looking at what age the biblical long-lived people are recorded as starting to sire children.
There are two genealogies (in chapters 5 and 11 of Genesis), plus some scattered statistics later on in the Bible, that give us tantalizing pieces of this information. In addition, there are two main versions of these genealogies that have come down to us in the Masoretic and Septuagint manuscripts, with differing numbers in the places where it matters to us most—the ages given when men fathered their first sons. For instance, did Arphaxad, grandson of Noah, really have his first son when he was only 35 years old, as the Masoretic says (Genesis 11:12)? Since the Masoretic says he lived a total of 438 years (Genesis 11:13), he would have had his first son when he was only 8% into his total lifespan. But if he was actually 135 when he fathered this son, according to the Septuagint he would have been 24% into his given lifespan of 565 years (Genesis 11:12,13), a more normal proportion throughout history, and even today. (The Masoretic and Septuagint manuscripts give different figures for Arphaxad’s total lifespan.)
The concept that the length of time between birth and sexual maturation might be related to the overall lifespan of humans has not been widely considered, and more work should be done in this area. It is this author’s suggestion that Cuozzo’s Neanderthal work points to the longer ages to fatherhood that are indicated in the Septuagint numerical genealogies, thus lengthening the earth’s historical timeline by some centuries.13
One way of getting around the problem of needing more time to account for historical events has been to accept the Masoretic figures, but to allow for time gaps in the early Genesis genealogies by claiming that the real amount of historical time was not intended to be known.14 This author rejects the idea of gaps in the Genesis 5 and 11 genealogies, on the basis that the numbers indicate that those genealogies are obviously intended to give a continuous time history.15
Cuozzo (1998a, pp. 253–4) prefers to keep to the Masoretic chronology while at the same time postulating a very slow rate of Neanderthal maturation; however, he admits that he has not studied this timeline angle (Cuozzo 2009, personal communication).
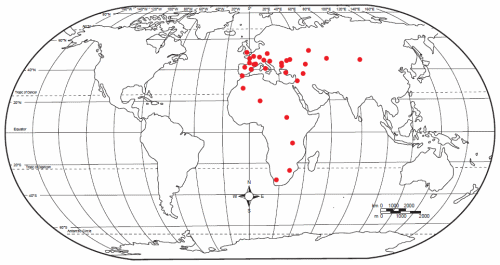
Fig. 8. World map showing spread of Neanderthal sites. Information for this map was taken by the author from various sources cited in the text of this paper, and is not intended to show all Neanderthal sites.
Implications of Distribution of Worldwide Neanderthal Sites
The widely held creationist belief that the Neanderthals must have been one of a number of family groups that traveled outwards in all directions from Babel worked fine as long as most of the discovered Neanderthal sites were in Europe and the Middle East (see map at List of Neanderthal sites 2009). This made sense, because it looked as if the Neanderthals were a nomadic family tribe that had headed northwest from Babel, split into groups, and settled in various places (Oard 1996, see map p. 58).
But the Neanderthal territory kept expanding (see fig. 8). Neanderthals have been found as far east as southern Siberia, close to China (Anitei 2007; Krause et al. 2007b), and down to the southern tip of Africa.16 Obviously they got around a lot more than had been previously thought.17 We see then that the very wide distribution of Neanderthals, as shown in Fig. 8, backs the idea that they were the people who dispersed in all directions from Babel, as Cuozzo believes. This also makes the Lubenow view of Neanderthals as a small variant group less likely.
A further look at Fig. 8 shows not only where Neanderthals have been found, but also where they have not been found. There are no Neanderthal sites in North or South America, Greenland or Australia. The question is, why not?
The world map (fig. 8) shows Neanderthal occupation along the western coast of Europe and northern Africa, meaning that ancient long-lived people migrated westward only that far. The most obvious barrier to further westward migration would have been the Atlantic ocean, with the corollary that the original land mass must have broken up some time before these people got to the western coasts.18 This would put the latest possible time of earth’s breakup fairly soon after the Babel dispersion; otherwise there would have been time for the early post-Flood people to have migrated westward to the territory that is now North America, and this apparently did not happen. (Obviously, if ever Neanderthals were found in North America, this would change the history of this continent. However, according to this map, it seems unlikely.)
Naturally, this matter is of no concern to those who believe that the earth broke up during the Flood. However, there are those who believe that the breakup of the earth occurred at the time of the birth of Peleg, because of the statement in Genesis 10:25 that “in his days was the earth divided.” This latter group has proposed various scenarios, with differing times when they think the earth broke up. Northrup (1996) and Lanser (2009) believe the breakup occurred 200 years after Babel, while Setterfield (1999) puts breakup 159 years after Babel. These periods of time would seem to be too long, as the probability is high that the Neanderthals could have arrived at and beyond the western shores of Europe from Babel in less than that time; however, this is difficult to prove one way or the other. Nelson does not propose an actual period of time between Babel and the subsequent earth breakup, but believes that the breakup of the earth helped to disperse people around the world, including the Americas (Nelson 2007, p. 102). The actual pattern of the Neanderthal sites refutes this, because the earth’s breakup had to have prevented, rather than caused, worldwide dispersal of peoples after Babel.
Based on Cuozzo’s work, we can say that people did not arrive in North America until after the human lifespan had shortened to the point that humans no longer developed the Neanderthal characteristics. This is in keeping with the disappearance of the Neanderthals everywhere before the end of the Ice Age, and the appearance of humans in North America in post-Ice-Age times. Not surprisingly, secular scientists are happy to put figures on these events for us: although they differ somewhat on when the earliest people arrived in North America, they are agreed that it was probably between 10,000 and 15,000 years ago on their evolutionary time scale. (For earliest human indications found in North America so far, see Gilbert et al. 2008). Since Neanderthals are commonly believed to have all disappeared by about 30,000 years ago (Lewin 1999, p. 156), this puts a gap between the demise of the Neanderthals elsewhere in the world and the arrival of humans in North America.19
Oard (2006a, see map p. 128) suggests that after the Babel dispersion people eventually migrated all across Asia, and crossed from the eastern point of Siberia to what is now Alaska, spreading south from there into the Americas. If we use our Neanderthals as a marker, the crossing to Alaska would most likely have taken place after the Ice Age, when the Neanderthals had “died out,” and could not have occurred early in the Ice Age as suggested by Oard (2000; 2006a, p. 130). Schroeder’s (2005) somewhat novel idea of immediate post-Babel people sailing around an icefree Arctic ocean, and migrating south through the Bering Strait to settle in countries around the Pacific Ocean could be supported only if Neanderthals could be found in these areas. As we see from Fig. 8, none have been found in these places so far; it appears that people headed out from Babel in all directions by land. (In any case, it would be questionable whether any of these small migrating groups, struggling for survival in an Ice-Age climate, would have had the resources to build a seafaring boat.) It was only later, in post-Neanderthal times, that people migrated to the Pacific rim lands, by which time the northern sea would have been full of ice (Oard 2006a, p. 96).
As a side comment, the huge territory that was occupied by Neanderthals has also had implications for those who advocate illness or climate as the cause of the Neanderthal morphology. It seems highly unlikely that all the widespread Neanderthals could have had rickets, syphilis, or whatever; or that the climate variations in these widespread regions would have affected all these people the same way.
The Ice Age, Geology, and Archaeology
As noted earlier, scholars believe that the Neanderthals “disappeared” some time before the end of the Ice Age. The Neanderthal demise and the Ice-Age meltdown are therefore historical events that occurred fairly close together in time.
Creationists understand this Ice Age to have occurred in post-Flood times. How many years in the past the Flood took place is subject to timeline differences of opinion (for example, Masoretic versus Septuagint genealogy numbers, and differing calculations of chronology of biblical events). In addition, there are assumptions necessarily inherent in calculations of the length of the Ice Age itself; Oard (2006a) goes into these in some detail. Indeed, if we attach the “disappearance” of the Neanderthals to the shortening of the human lifespan, the actual time in history when people no longer displayed Neanderthal characteristics may have differed in various parts of the world because of lifestyle, climate, or other factors. The Ice-Age meltdown, therefore, can only provide us with a general historical marker with respect to the Neanderthals.
It is hard for us to imagine what the world must have been like during the catastrophic period of the melting of the great continental ice sheets and the glaciers in high mountains. Enormous amounts of water were released from the melting ice, causing flooding and great changes in the landscape, and raising world ocean levels from their minimum at the height of the glaciation to approximately their level today (Lamb 1997, pp. 114–116).
One country that has been affected quite clearly by this kind of geologic history is Egypt. The high mountains in the African countries south of Egypt developed large glaciers during the Ice Age; in the time of the Ice-Age melting, vast amounts of water flowed down the Nile tributaries and northward in the Nile valley. At about the same time, a belt of heavy monsoon rains moved northward in Africa, adding to the extraordinary Nile inundations of this time (Bard 1999, p. 16; Close 1996, pp. 43–46). Great amounts of silt washed down the Nile along with all this torrential water and largely formed the huge fan-shaped delta at Egypt’s north end on the Mediterranean (Stanley and Warne 1993). This catastrophic flooding effectively destroyed many signs of prehistoric Ice-Age habitation along the Nile, of which only remnants have been found by archaeologists (Close 1996, p. 43; Edwards 1970, pp. 62–72; Mithen 2003, pp. 451–452; Shaw 2002, p. 29). We might expect that the very earliest people who lived in Egypt before the Ice-Age meltdown would have been Neanderthals who had migrated from Babel, and whose descendants moved elsewhere when the meltdown flooding began. During this period of the inundations, Egypt was effectively unpopulated. After the era of Ice-Age meltdown flooding ended, Egypt was eventually repopulated by modern people; therefore all known civilization in Egypt from earliest predynastic times on is post Ice Age. This sequence of events in earliest ancient Egypt has been well developed by various historians (see, for example, Midant-Reynes, 2000; Shaw 2002).
Parenthetically, there are wider implications of this early Egyptian history. We know that, to avoid famine in Canaan, Abraham picked up and went to Egypt (Genesis 12:10–20). There was a pharaoh reigning there then, a pharaoh that was powerful enough to intimidate Abraham, who lied about Sarah’s marital status to protect himself (Genesis 12:13). This means that by the time of Abraham’s visit, Egypt’s population and civilization had developed sufficiently to have a strong ruler in place, and the Ice Age was long past. The currently accepted 700-year Ice Age (Oard 2006b) does not fit into the 427 years allowed by Usshur (1658, p. 25) and Jones (2007, CD and p. 57) for all the geological and historical events that occurred between the Flood and Abraham’s Egyptian sojourn. Furthermore, according to the Usshur/Jones chronology, a 700-year Ice Age starting after the Flood would place Jacob’s arrival in Egypt (Genesis 46, 47) during the period of the Nile inundations from the Ice-Age meltdown, while the delta, where Goshen was located, was still being formed from the silt. One possible solution to this problem would be to shorten the complete Ice Age, meltdown and all, to about 300 years. Another would be to go to the longer timeline offered by the Septuagint chronology, as discussed earlier. What is clear is that the current state of acceptance of both the Masoretic genealogies and the 700-year Ice Age is incompatible with the biblical account and Egyptian history.
Another place that yields historical and geological information from the Ice Age is the large cave at Shanidar in the Zagros mountains of northwest Iraq, where a number of Neanderthals have been found (Hirst 2009; Trinkaus and Shipman 1993, pp.334–341.) Because of the Neanderthal occupation, we know that this cave had to have been there during at least part of the Ice Age, and also had to have survived the end-of-Ice-Age flooding which took place in the area (the cave is located on a hillside high above the valley bottom). In addition, we know that the mountains in this area must have been already formed well before the end of the Ice Age.
As in Egypt, so in Mesopotamia the Ice-Age meltdown made an enormous difference in the geology of the country. When the Shanidar Neanderthals looked out of their cave toward the Greater Zab River valley of Northern Iraq, they could not have envisioned the great torrents of water that would later flow down this tributary to the Tigris River at the end of the Ice Age. The entire southern half of Mesopotamia is composed of the delta formed largely by silts washed down the Tigris and Euphrates rivers from the north during the great floods of the Ice-Age meltdown (see, for instance, McIntosh 2005, pp. 8–9; Nutzel 1979; Persian Gulf Once Dry, Green, and Inhabited by Humans 2007; Postgate 1992). This has implications for those who look for the Tower of Babel on this delta, since everything in southern Mesopotamia has to be post Ice Age, including the ancient city of Babylon.
At the peak of the Ice Age, world ocean levels were at their minimum because of the huge amount of water that had been frozen onto the thick continental ice sheets.20 The ancient stone constructions around the world that are located under water, not far from current shorelines, may well have been built during the time of the lower ocean levels of the Ice Age. Secular writers are understandably mystified by these sophisticated prehistoric constructions that do not coincide with an evolutionary worldview (Hancock 2002); one might wonder whether these underwater cities were built by Ice-Age people whom we would define as Neanderthals. A Neanderthal skull fragment has been recently dredged up from the North Sea (Vieru 2009); since this sea has an average depth of only 95 m, it would most likely have been dry land at the peak of the Ice Age, and this Neanderthal could well have lived there at that time (About the North Sea 2009).
In a somewhat novel view of history, von Fange (1994, see chart p. 222) places the entire Ice Age and its meltdown before the Babel dispersion. This sequence of events does not fit with known facts for a number of reasons, one of which is that Neanderthals (that is, mankind) had already scattered widely because of the Babel dispersion before the end of the Ice Age. It is beyond the scope of this paper to go more deeply into other aspects of why the Ice Age could not have ended before Babel.
Additionally, the controversial question of whether human tracks appear together with dinosaur tracks is affected by this discussion of Neanderthals and the Ice Age. In North America, especially, the subject is somewhat mixed up with disagreement as to whether certain tracks are actually human or not,21 and whether or not track layers are post Flood.22 What we can say is that if large dinosaur tracks in the Americas or Australia are post Flood, any human-looking tracks that appear with them cannot have been made by humans. This is because the large dinosaurs died out long before post-Ice-Age modern humans reached these lands23; only in countries where Neanderthal sites occur could post-Flood dinosaurs and humans have coexisted.
Closing comments
The question is necessarily raised whether traditional paleoanthropology as a science has been adequate for study of Neanderthals without the additional input of specialized medical expertise. Where only traditional paleoanthropology has been applied, not only has the subject of the Neanderthals’ great age at death not even been considered, but faulty medical notions have been accepted (for example, with respect to disease as a supposed cause of the distinctive Neanderthal characteristics). The result has been that application of paleoanthropology alone, without medical input, has led creationists down quite a different path of thinking than the one they might have taken if orthodontic and other applicable medical expertise had been included.
This paper also points out the importance of harmonization of various areas of scientific knowledge with respect to any study. In this discussion of the Neanderthals, we see that paleoanthropology, medicine (including orthodontics, skeletal studies, pathology, ageing studies), computer simulation, genetics, history, paleoarchaeology, geography and geology have all come into play, in addition to the biblical record. It should always be understood that one area of scientific knowledge does not supersede the others in importance. All scientific disciplines involved must be considered and all must contribute equally; otherwise the conclusions may be skewed, and may even be incorrect.
On the matter of the Neanderthals, there is no compromise ground between the two sets of conclusions that have been reached by the two major creationist authorities (Cuozzo and Lubenow), because those conclusions are largely mutually exclusive. Moreover, the implications of this divergence of thought are enormous for the young-earth creationist model, as has been shown in this paper.
The Neanderthals have a lot to say to us, if we will listen. But to hear what the Neanderthals are saying, we must consider the ramifications of Cuozzo’s work which, in turn, balances on whether his scientific expertise is valid or not. This leads to the inevitable admission that if we cannot falsify Cuozzo’s scientific conclusions on the Neanderthals, we are forced to accept them. Ignoring them is not an honest scientific option.
Acknowledgment
I would like to thank Dr. Jack Cuozzo for his patience and generosity in answering my many questions during the time of writing this paper.
References
About the North Sea: Key Facts. 2009. Safety at Sea Newsletter. Retrieved Oct. 30, 2009, from http://www.safetyatsea.se/index.php?section=northsea.
Acton, R. 1978. Bone disease simulating ancient age in prehuman fossils. Impact Series #59, May 1, 1978. San Diego, California: Institute for Creation Research.
Albert, A. M., K. Ricanek, and E. Patterson. 2007. A review of the literature on the aging adult skull and face: Implications for forensic science research and applications. Journal of Forensic Science International 172, no. 1:1–9. Retrieved Oct. 30, 2009 from http://people.uncw.edu/pattersone/research/publications/AlbertRicanekPatterson_FSI2007.pdf.
Anderson, D. 2006. A new Neandertal/modern human fossil hybrid? Creation Ministries International featured article online. November 14, 2006. Retrieved August 28, 2009, from http://creation.com/a-new-neandertal-modern-humanfossil-hybrid.
ANI. 2009. Neanderthals may have gone extinct because of their small population size. Thaindian News, July 17, 2009. Retrieved August 31, 2009, from http://www.thaindian.com/newsportal/health/neanderthals-may-have-gone-extinctbecause-of-their-small-population-size_100219091.html.
Anitei, S. 2007. Neanderthals reached China! Fossils of Neanderthals from southern Siberia. Softpedia, October 1, 2007. Retrieved August 30, 2009, from http://news.softpedia.com/newsPDF/Neanderthals-Reached-China-67178.pdf.
Argue, D., M. Morwood, T. Sutikna, Jatmiko, and W. Saptomo. 2009. Homo floresiensis: A cladistic analysis. Journal of Evolution, pub. online July, 2009. doi:10.1016/j.jhevol.2009.05.002.
Ashcraft, C. W. 2004. Genetic variability by design. Technical Journal 18, no. 2:98–104.
Baab, K. L. and K. P. McNulty. 2008. Size, shape, and asymmetry in fossil hominins: The status of the LB1 cranium based on 3D morphometric analyses. Journal of Human Evolution. Online preview article retrieved Aug. 13, 2009, from http://www.karenbaab.com/uploads/1/5/6/4/1564155/baabmcnulty2008_size_scaling_asymmetry_lb1.pdf.
Banks, W. E. F. d’Errico1, A. Townsend Peterson, M. Kageyama, A. Sima, and M.-F. Sánchez-Goñi. 2008. Neanderthal extinction by competitive exclusion. PLoS ONE 3, no. 12:e3972. Retrieved Aug. 18, 2009, from http://www.plosone.org/article/info:doi/10.1371/journal.pone.0003972#top.
Bard, K. A. 1999. Encyclopedia of the archaeology of ancient Egypt. London: Routledge.
Bartlett, S. P., R. Grossman, and L. A. Whitaker. 1992. Age-related changes of the craniofacial skeleton; An anthropometric and histologic analysis. Plastic and Reconstructive Surgery: Journal of the American Society of Plastic Surgeons 90, no. 4:592–600.
Beasley, G. J. 1992. A possible creationist interpretation of archaic fossil human remains. Technical Journal 6, no. 2:138–167.
Behrents, R. 1985. Growth in the aging craniofacial skeleton, Monograph 17. Center for Growth and Development. Ann Arbor, Michigan: University of Michigan.
Bowden, M. 1981. Ape-Man: Fact or Fallacy? 2nd ed. Bromley, Kent, England: Sovereign Publications.
Bowden, M. 1988. Homo erectus—a fabricated class of ‘apemen.’ Ex Nihilo Technical Journal 3, no. 1:152–153.
Briggs, A. W. et al. 2009. Targeted retrieval and analysis of five Neandertal mtDNA genomes. Science 325. no. 5938: 318–321. Retrieved abstract August 10, 2009 from http://www.sciencemag.org/cgi/content/abstract/325/5938/318.
Brown, P. 1997. Kow Swamp: Is it Homo erectus? Retrieved May 8, 2009 from http://www.talkorigins.org/faqs/homs/kowswamp.html.
Brown, P., T. Sutikna, M. J. Morwood, R. P. Soejono, Jatmiko, E. W. Saptomo, and R. A. Due. 2004. A new small-bodied hominin from the Late Pleistocene of Flores, Indonesia. Nature 431:1055–1061.
Brow ridge. n.d. The Museum for Comparative Anthropogeny, web site of the Center for Academic Research & Training in Anthropogeny. Retrieved January 1, 2010 from http://carta.anthropogeny.org/moca/topics/brow-ridge.
Carnieri, E. 2006. Who killed the Neanderthals? Human Evolution 21:337–340.
Carroll, L. 1865. Alice’s Adventures in Wonderland. Oxford: MacMillan 1865, and New York: Appleton 1866.
Carter, R. W. 2009. The Neanderthal mitochondrial genome does not support evolution. Journal of Creation 23. no. 1: 40–43.
Close, A. E. 1996. Plus ca change: The Pleistocene–Holocene transition in Northeast Africa. In Humans at the end of the Ice Age: The archaeology of the Pleistocene, ed. L. G. Straus. New York: Plenum Pub. Corp.
Conrad, E. 1986. Creationists and Neandertal. Creation Evolution Journal 6, no. 3:24–33.
Criswell, D. 2009. Neanderthal DNA and modern humans. Creation Research Society Quarterly 45, no. 4:246–254.
Cuozzo, J. W. 1987. Earlier orthodontic intervention: A view from prehistory. The Journal of the New Jersey Dental Association 58, no. 4:33–40.
Cuozzo, J. W. 1991. Asymmetric adult growth and mechanical advantage in the etiology of the myofacial pain dysfunction syndrome: A case report. The Journal of the New Jersey Dental Association 62, no. 4:23–29, 31.
Cuozzo, J. W. 1994. Neanderthal children’s fossils: Reconstruction and interpretation distorted by assumptions. Creation Ex Nihilo Technical Journal 8, no. 2:166–178. Retrieved October 30, 2009 from http://creation.com/neandertal-childrens-fossils.
Cuozzo, J. W. 1998a. Buried alive. Green Forest, Arkansas: Master Books.
Cuozzo, J. W. 1998b. What happens to the craniofacial structure of humans who live past 100 years? Neanderthal similarities. In Proceedings of the fourth international conference on creationism, ed. R. E. Walsh, pp. 103–119. Pittsburgh, Pennsylvania: Creation Science Fellowship.
Cuozzo, J. W. 2003. Buried alive supplement CD.
Cuozzo, J. W. 2009. Neanderthal DNA. Retrieved September 10, 2009 from http://jackcuozzo.com/dna.html.
Currat, M. and L. Excoffier. 2004. Modern humans did not admix with Neanderthals during their range expansion into Europe. PLoS Biology 2, no. 12:e421. doi:10.1371/journal.pbio.0020421.
Custance, A. C. 1968. Fossil man in the light of the record in Genesis. Creation Research Society Quarterly 5. no. 1:5–22.
Dean, C. M. G. Leakey, D. Reid, F. Schrenk, G. T. Schwartz, C. Stringer, and A. Walker. 2001. Growth processes in teeth distinguish modern humans from Homo erectus and earlier hominins. Nature 414:628–631.
Dobson, J. E. 1998. The iodine factor in health and evolution. The Geographical Review 88:1–28.
Drell, R. R. 2000. Neanderthals: A history of interpretation. Oxford Journal of Archaeology 19(1):1–24. Retrieved July 25, 2009, from http://archaeology.about.com/gi/dynamic/offsite.htm?zi=1/XJ&sdn=archaeology&cdn=education&tm=36&f=00&su=p897.4.336.ip_&tt=2&bt=1&bts=1&zu=http%3A//dx.doi.org/10.1111/1468-0092.00096.
Edwards, I. E. S. 1970. The Cambridge ancient history: Part I, 3rd ed. Cambridge, England: Cambridge University Press.
Fabre, V., S. Condemi, and A. Degioanni. 2009. Genetic evidence of geographical groups among Neanderthals. PLoS ONE 4. no. 4:e5151. Retrieved August 13, 2009 from http://www.plosone.org/article/info:doi/10.1371/journal.pone.0005151.
Field, E. 1894. The duel. In Love-Songs of Childhood. New York: Scribner.
Flores, J. C. 1998. A mathematical model for Neanderthal extinction. Journal of Theoretical Biology 191, no. 3: 295–298.
Foley, J. 2008. Comparison of all skulls. Retrieved August 4, 2009, from Talk Origins online: http://www.talkorigins.org/faqs/homs/compare.html.
Freeman, T. R. 2005. The Genesis 5 and 11 fluidity question. Journal of Creation 19, no. 2:83–90.
Garton, M. 1996. The pattern of fossil tracks in the geological record. Creation Ex Nihilo Technical Journal 10, no. 1: 82–100.
Gilbert, M. T. P., et al. 2008. DNA from pre-Clovis human coprolites in Oregon, North America. Science 320. no. 5877:786–789. DOI: 10.1126/science.1154116.
Gish, D. T. 1985. Evolution: The challenge of the fossil record. El Cajon, California: Creation-Life Publishers.
Gornitz, V. 2007. Sea level rise, after the ice melted and today. NASA: Goddard Institute for Space Studies. Retrieved Oct. 25, 2009 from: http://www.giss.nasa.gov/research/briefs/gornitz_09/.
Green, R. E. et al. 2008. A complete Neandertal mitochondrial genome sequence determined by high-throughput sequencing. Cell 134, no. 3:416–426.
Guagliardo, M. F. 1982. Craniofacial structure, aging, and dental functions: Their relationships in adult human skeletal series. Doctoral dissertation. Knoxville, Tennessee: University of Tennessee.
Gugliotta, G. 1999. Geographer suggests Neanderthals were just cretins. The Japan Times, June 1, 1999. Retrieved Septe,ber 23, 2009 from http://www.trussel.com/prehist/news125.htm.
Hancock, G. 2002. Underworld: The mysterious origins of civilization. New York: Three Rivers Press.
Hirst, K. K. 2009. Shanidar Cave (Iraq). Retrieved November 11, 2009 from http://archaeology.about.com/od/shthroughsiterms/g/shanidar.htm.
Hodge, B. 2007. How old is the earth? Retrieved November 4, 2009 from http://www.answersingenesis.org/articles/2007/05/30/how-old-is-earth.
Hoffecker, J. F. 2002. Desolate landscapes: Ice-age settlement in Eastern Europe. New Brunswick, New Jersey: Rutgers University Press.
Hoffecker, J. F. 2005. A prehistory of the north: human settlement of the higher latitudes. New Brunswick, New Jersey: Rutgers University Press.
Homo erectus. 2009a. In Encyclopædia Britannica. Retrieved August 10, 2009 from http://www.britannica.com/EBchecked/topic/270386/Homo-erectus/.
Homo erectus. 2009b. In Wikipedia. Retrieved November 16, 2009, from http://en.wikipedia.org/wiki/Homo_erectus.
Horan, R. D., E. Bulte, and J. F. Shogren. 2005. How trade saved humanity from biological exclusion: An economic theory of Neanderthal extinction. Journal of Economic Behavior & Organization 58, no. 1:1–29.
Howell, F. C. 1957. The evolutionary significance of variation and varieties of “Neanderthal” man. The Quarterly Review of Biology 32. no. 4:330–347.
Howell, F. C. 1967. The hominization process. In Human Evolution: Readings in physical anthropology, ed. N. Korn. New York: Rinehart and Winston.
Human stabbed a Neanderthal, evidence suggests. 2009. Answers in Genesis, News to note, July 25, 2009. Retrieved October 30, 2009 from http://www.answersingenesis.org/articles/2009/07/25/news-tonote-07252009.
Israel, H. 1977. The dichotomous pattern of craniofacial expansion during aging. American Journal of Physical Anthropology 47, no. 1:47–51.
Ivanhoe, F. 1970. Was Virchow right about Neandertal? Nature 227:577–579.
Jaroncyk, R. 2007. A new Neandertal/modern human fossil hybrid? Journal of Creation 21, no. 1:7–8.
Jelinek, A. J. 1982. The Tabun Cave and Paleolithic man in the Levant. Science 216:1369–1375.
Jimenez-Espejo, F. J., F. Martínez-Ruiz, C. Finlayson, A. Paytane, T. Sakamoto, M. Ortega-Huertas, G. Finlayson, K. Iijima, D. Gallego-Torres and D. Fac. 2007.Climate forcing and Neanderthal extinction in Southern Iberia: Insights from a multiproxy marine record. Quaternary Science Reviews 26, no. 7–8:836–852.
Johnson, W. 1982. The crumbling theory of evolution. The Gap, Brisbane,Australia: published by author.
Jones, D-W. 1998. How old is that fossil (in the layer)? Retrieved May 8, 2009 from http://www.ridgecrest.ca.us/~do_while/sage/v3i2f.htm.
Jones, F. N. 2007. The chronology of the Old Testament, 16th ed. Green Forest, Arkansas: Master Books.
Josephus, F. 1988. The works of Josephus. W. Whiston, trans. Peabody, Massachusetts: Hendrickson Publishers.
Jue, D. S. 1990. Cranial capacity and endocranial casts. Creation Ex Nihilo Technical Journal 4, no. 1:56–65.
Klein, R. G. 1989. The human career: Human biological and cultural origins. Chicago: University of Chicago Press.
Klein, R. G. 2003. Paleoanthropology: Whither the Neanderthals? Science 299, no. 5612:1525–1527. DOI: 10.1126/science.1082025.
Krause, J. et al. 2007a. The derived FOXP2 variant of modern humans was shared with Neandertals. Current Biology 17. no. 21:1908–1912.
Krause, J. et al. 2007b. Neanderthals in central Asia and Siberia. Nature 449:902–904.
Krogman, W. M. 1972. Child Growth. Ann Arbor, Michigan: University of Michigan Press.
Kuban, G. J. 2009. The Paluxy dinosaur/”man track” controversy. Retrieved November 1, 2009 from http://paleo.cc/paluxy/paluxy.htm.
Kuhn , S. L. and M. C. Stiner. 2006. What’s a mother to do? The division of labor among Neandertals and modern humans in Eurasia. Current Anthropology 47(6):953–980.
Lahr, M. M. 1996. The evolution of modern human diversity; A study of cranial variation. Cambridge University Press.
Lamb, H. H. 1997. Climate, history and the modern world. London: Routledge.
Lanser, R. D. 2009. Making sense of the days of Peleg. Bible and Spade 22, no. 2:51–64.
Larson, S. G. et al. 2007. Homo floresiensis and the evolution of the hominin shoulder. Journal of Human Evolution 53. no. 6:718–731.
Leakey, R. 1996. The origin of humankind. New York: Basic Books.
Lewin, R. 1999. The origin of modern humans. 4th ed. United Kingdom: Blackwell Science.
Life expectancy at birth. 2008. The world factbook. Directorate of Intelligence of the U.S. Central Intelligence Agency.
Line, P. 2005. Fossil evidence for alleged apemen—Part 1: The genus Homo. Journal of Creation 19, no. 1:22–32.
Line, P. 2007. Inconvenient Neandertaloids. Technical Journal 21, no. 1:15–19.
List of Neanderthal sites. 2009. Wikipedia. Retrieved January 1, 2010 from http://en.wikipedia.org/wiki/List_of_Neanderthal_sites.
Lovgren, S. 2005. Chimps, humans 96 percent the same, gene study finds. National Geographic News. Retrieved November 25, 2009 from http://news.nationalgeographic.com/news/2005/08/0831_050831_chimp_genes.html.
Lubenow, M. L. 1998. Recovery of Neandertal mtDNA: An evaluation. CEN Technical Journal 12, no. 1:87–97.
Lubenow, M. L. 2000. Lagar Velho 1 child skeleton: a Neanderthal/modern human hybrid. Technical Journal 14, no. 2:6–8. Retrieved October 8, 2009 from http://creation.com/images/pdfs/tj/tjv14n2_neandertal.pdf.
Lubenow, M. L. 2004. Bones of contention: A creationist assessment of human fossils, 2nd ed. Grand Rapids, Michigan: Baker.
Lubenow, M. L. 2006. The Neanderthals: Our worthy ancestors. Part I: The DNA evidence. Retrieved October 30, 2009 from http://www.answersingenesis.org/articles/am/v1/n2/worthy-ancestors.
Lubenow, M.L. 2007. The Neanderthals: Our worthy ancestors. Part II: The fossil and archaeological evidence. Retrieved October 30, 2009, from http://www.answersingenesis.org/articles/am/v2/n1/worthy-ancestors-2.
McIntosh, J. 2005. Ancient Mesopotamia: New perspectives. Santa Barbara, California: ABC-CLIO.
McKie, R. 2009. How Neanderthals met a grisly fate: devoured by humans. The Observer, Sunday 17 May 2009. Retrieved September 1, 2009 from http://www.guardian.co.uk/science/2009/may/17/neanderthals-cannibalismanthropological-sciences-journal.
Mehlert, A. W. 1994. Homo erectus ‘to’ modern man: Evolution or human variability? Journal of Creation 8, no. 1:105–116.
Mellars, P. 1996. The Neanderthal legacy: An archaeological perspective from western Europe. Princeton, New Jersey: Princeton University Press.
Midant-Reynes, B. 2000. The prehistory of Egypt from the first Egyptians to the first pharaohs. Oxford, England: Blackwell; Malden, Massachusetts: Blackwell.
Miller, M. 2001. The Neanderthal demise: was it love, or was it war (with modern humans)? Athena Review 2, no. 4. Retrieved October 29, 2009 from http://www.athenapub.com/8miller1.htm.
Milner, C. S., R. A. Neave, and C. M. Wilkinson. 2001. Predicting growth in the aging craniofacial skeleton. Paper presented at the 9th Biennial Meeting of the International Association for Craniofacial Identification, FBI, Washington, DC, July 2000. Retrieved October 30, 2009 from http://www.fbi.gov/hq/lab/fsc/backissu/july2001/milner.htm.
Mithen, S. 2003. After the ice. Cambridge, Massachusetts: Harvard University Press.
Morgan, J. 2009. Neanderthals ‘distinct from us.’ BBC News, Chicago. 12 February 2009. Retrieved August 28, 2009 from http://news.bbc.co.uk/2/hi/health/7886477.stm.
Morris, H. M. 1976. The Genesis Record. Grand Rapids, Michigan: Baker Books.
Moskowitz, C. 2008. Neanderthal brains grew like ours. Livescience web site, September 8, 2008. Retrieved September 24, 2009 from http://www.livescience.com/history/080908-neanderthal-skulls.html.
Murdock, M. 2004. Sahelanthropus tchadensis—the ambiguous ape. Technical Journal 18, no. 3:110–115.
Murdock, M. 2006. These apes were made for walking: The pelves of Australopithecus afarensis and Australopithecus africanus. Technical Journal 20, no. 2:104–112.
Neanderthal. 2009. Wikipedia. Figs 1, 2, 3, 4, and 7 retrieved November 16, 2009 from http://en.wikipedia.org/wiki/ Neanderthal. In addition, the courtesy of the following link is requested for fig. 7: http://www.ifi.uzh.ch/staff/zolli/CAP/Main_face.htm.
Nelson, J. W. 2004. Genetic variability and human history. Technical Journal 18, no. 3:18–23.
Nelson, D. P. 2007. Peleg: Early earth movements. Clarksburg, West Virginia: Pelegpress.
Neuroscience and intelligence. 2009. Wikipedia. Modified September 29, 2009. Retrieved October 5, 2009, from http://en.wikipedia.org/wiki/Neuroscience_and_intelligence.
Northrup, B. E. 1996. On finding an Ice Age book. Published online. Retrieved September 18, 2009, at http://ldolphin.org/iceage.html.
Oard, M. 1996. Life in the great ice age. Green Forest, Arkansas: Master Books.
Oard, M. 2000. Did ‘old world early Paleolithic’ people travel to North America? Journal of Creation 14, no. 3:3–5.
Oard, M. 2003a. Neandertal man—the changing picture. An overview of how this alleged ‘subhuman’ is being progressively rehabilitated, despite the evolutionary bias resisting the trend. Creation 25, no. 4:10–14.
Oard, M. 2003b. Do genetic differences disprove that Neandertals and modern humans interbred? Journal of Creation 17, no. 2:18.
Oard, M. 2003c. In the footsteps of giants. Creation 25, no. 2:10–12.
Oard, M. 2005. Book review: Bones of contention—revised edition of 1992 book by Lubenow. Creation Research Society Quarterly 42, no. 1:79.
Oard, M. 2006a. Frozen in time. Green Forest, Arkansas: Master Books.
Oard, M. 2006b. Where does the Ice Age fit? In The new answers book, ed. K. Ham, pp. 207–219. Green Forest, Arkansas: New Leaf Press. Retrieved Oct. 26, 2009, from http://www.answersingenesis.org/articles/nab/where-does-ice-age-fit.
Oard, M. 2009. New footprints from Ileret, Kenya. Journal of Creation 23, no. 2:3–4.
O’Connor, C. F., R. G. Franciscus, and N. E. Holton. 2004. Bite force production capability and efficiency in Neandertals and modern humans. American Journal of Physical Anthropology 127, no. 2:129–151.
Occam’s razor. 2009. Wikipedia. Retrieved August 28, 2009 from http://en.wikipedia.org/wiki/Occam%27s_razor.
Parker, G. 2006. The fossil evidence. Creation: facts of life. Green Forest, Arkansas: Master Books. Retrieved October 31, 2009 from 3.6 Human Beings.
Paul, G.S. (ed.). 2000. The Scientific American book of dinosaurs. New York, New York: St. Martin’s Press.
Persian Gulf Once Dry, Green, and Inhabited by Humans 2007. Suburban Emergency Management Project. Biot Report #422: May 15, 2007. Retrieved November 27, 2009, from http://www.semp.us/publications/biot_reader.php?BiotID=422.
Phillips, D. 2000. Neanderthals are still human! Retrieved July 23, 2009 from http://www.icr.org/article/neanderthalsare-still-human/.
Ponce de Leon, M. S. et al. 2008. Neanderthal brain size at birth provides insights into the evolution of human life history. In Proceedings of the National Academy of Sciences of the United States of America 105, no. 37:13764–68.
Postgate, J. N. 1992. Early Mesopotamia: Society and economy at the dawn of history. London, England: Routledge.
Purdom, G. 2007. Red-haired, fast-talking Neanderthals. Answers in Depth 2:163–165. Nov. 21, 2007, Retrieved October 31, 2009, from http://www.answersingenesis.org/articles/aid/v2/n1/red-haired-fast-talking.
Rae, T. C., U. S. Viðarsdóttir, N. Jeffery and A. T. Steegmann, Jr. 2006. Developmental response to cold stress in cranial morphology of Rattus; Implications for the interpretation of climatic adaptation in fossil hominins. Proceedings of the Royal Society: Biological Sciences 273, no. 1601:2605–2610.
Rana, F. and H. Ross. 2005. Who was Adam? A creation model approach to the origin of man. Colorado Springs, Colorado: NavPress.
Reed, J. K., A. S. Kulikovsky, and M. J. Oard. 2009. Can recolonization explain the rock record? Creation Research Society Quarterly 46, no. 1:27—39.
Reed, J. K. and M. J. Oard (eds.). 2006. The geologic column: Perspectives within diluvial geology. Chino Valley, Arizona: Creation Research Society Books.
Regal, B. 2004. Human evolution: a guide to the debates. Santa Barbara, California: ABC-CLIO.
Richards, M. P., K. Harvati, V. Grimes, C. Smith, T. Smith, J.-J. Hublin, P. Karkanas and E. Panagopoulou. 2008. Strontium isotope evidence of Neanderthal mobility at the site of Lakonis, Greece, using laser-ablation PIMMS. Journal of Archaeological Science 35, no. 5:1251–1256.
Robbins, A. 2009. Neanderthal life spans and the Bible. Bible and Spade 22, no. 2:46–50.
Robertson, M. and J. Sarfati. 2003. More evidence Neandertals were fully human. Technical Journal 17, no. 3:13.
Rosnau, P. O., J. Auldaney, G. F. Howe, and W. Waisgerber. 1989a. Are human and mammal tracks found together with the tracks of dinosaurs in the Kayenta of Arizona? Part 1. Creation Research Society Quarterly 26. no. 2: 41–48.
Rosnau, P. O., J. Auldaney, G. F. Howe, and W. Waisgerber. 1989b. Are human and mammal tracks found together with the tracks of dinosaurs in the Kayenta of Arizona? Part 2. Creation Research Society Quarterly 26. no. 3:77–99.
Rozzi, F. V. R. and de Castro, J. M. B. 2004. Surprisingly rapid growth in Neanderthals. Nature 428:936–939.
Rozzi, F. V. R,. F. V. Ramirez Rozzi, F. d’Errico, M. Vanhaeren, P. M. Grootes, B. Kerautret, and V. Dujardin. 2009. Cutmarked human remains bearing Neandertal features and modern human remains associated with the Aurignacian at Les Rois. Journal of Anthropological Sciences 87:153–185.
Rusch, W. H. 1964. Eskimos and Aleuts: Their origins and evolution. Creation Research Society Quarterly 1, no. 1: 13–14.
Rusch, W. H. 1971. Human footprints in rocks. Creation Research Society Quarterly 7, no. 4:201–213.
Sanders, R. 1999. Bone fragments link Neanderthals with cannibalism. Retrieved August 19, 2009 from http://www.berkeley.edu/news/berkeleyan/1999/1006/cannibals.html.
Sanford, J. C. 2008. Genetic entropy and the mystery of the genome. Waterloo, New York: FMS Publications.
Sarfati, J. 2004. Refuting compromise. Green Forest, Arkansas: Master Books.
Schrenk, F. and S. Muller. 2008. The Neanderthals, p. x. London: Taylor & Francis.
Schroeder, L. L. 2005. A possible post-Flood human migration route. Technical Journal 19, no. 1:65–72.
Schultz, M. 2008. Rudolf Virchow. Emerging Infectious Diseases 14, no. 9:1480–1481.
Setterfield, B. 1999. Rev. 2003 and 2007. Ancient chronology in Scripture. Retrieved September 19, 2009 from https://web.archive.org/web/20090506092501/http://www.setterfield.org/000docs/scriptchron.htm#toadam.
Shaw, I. (ed.). 2002. The Oxford illustrated history of ancient Egypt. Oxford, England: Oxford university Press.
Shaw, R. D. 1970. Fossil man: Ancestor or descendant of Adam? Creation Research Society Quarterly 6, no. 4:172–181.
Shea, J. J. 2001. Modern human origins and Neanderthal extinctions in the Levant. Athena Review 2, no. 4:21–32.
Silvestru, E. 2004. Human and dinosaur fossil footprints in the Upper Cretaceous of North America? Journal of Creation 18, no. 2:114–120.
Snelling, A. 2007. A catastrophic breakup. Answers 2, no. 2:44–48. Retrieved November 1, 2009 from http://www.answersingenesis.org/geology/plate-tectonics/a-catastrophic-breakup/.
Snelling, A. 2009. Radiometric dating 101: Part 2. Answers 4, no. 4:71–73.
Soficaru, A., A. Dobos, and E. Trinkaus. 2006. Early modern humans from the Pestera Muierii, Baia de Fier, Romania. Proceedings of the National Academy of Sciences 103, no. 46:17196–17201.
Sorensen, B. n.d. Demography and the extinction of the European Neanderthals. Retrieved September 16, 2009, from http://energy.ruc.dk/Neanderthal%20Demography.pdf.
Spencer, M. A. and B. Demes. 1993. Biomechanical analysis of masticatory system configuration in Neandertals and Inuits. American Journal of Physical Anthropology 91, no. 1:1–20.
Speth, J. D. 2004. News flash: Negative evidence convicts Neanderthals of gross mental incompetence. World Archaeology 36, no. 4:519–526. Debates in World Archaeology.
Stanley, D. J. and A. G. Warne. 1993. Sea level and initiation of predynastic culture in the Nile delta. Nature 363: 435–438.
Stone, T. and M. L. Cupper 2003. Last glacial maximum ages for robust humans at Kow Swamp, southern Australia. Journal of Human Evolution 45, no. 2:99–111.
Stringer, C. and R. McKie. 1996. African exodus: The origins of modern humanity. New York: Henry Holt.
Swaminathan, N. 2007. Cave speak: Did Neandertals talk? Discovery of the human variant of the FOXP2 gene in Neandertals suggests they may have had language skills. Retrieved September 6, 2007, from http://www.scientificamerican.com/article.cfm?id=cave-speak-didneandertal.
Tattersall, I. 2007. Neanderthals, Homo sapiens, and the question of species in paleoanthropology. Journal of Anthropological Sciences 85:139–146.
Taylor, I. T. 2008. In the minds of men: Darwin and the new world order, 6th ed. Foley, Minnesota: TFE Publishing in cooperation with Creation Moments.
Tinkle, W. J. 1968. The ancestry of man. Creation Research Society Quarterly 5, no. 1:42–45.
Trinkaus, E. 1978. Hard times among the Neanderthals. Natural History 87, no. 10:58–63.
Trinkaus, E. 2007. European early modern humans and the fate of the Neandertals. Proceedings of the National Academy of Sciences 104, no. 18:7367–7372.
Trinkaus, E. and P. Shipman. 1993. The Neanderthals: Changing the image of mankind. New York: Alfred A. Knopf.
Turkana Boy. 2009. Wikipedia. Retrieved August 6, 2009 from http://en.wikipedia.org/wiki/Turkana_Boy.
Twain, M. 1883. Life on the Mississippi. Boston, Massachusetts: James R. Osgood & Co.
Tyler, D. J. 2005. The amazing shrinking humans from Flores. Origins Journal 41:8–11.
Tyler, D. J. 2006. Recolonisation and the Mabbul. In The geologic column: Perspectives within diluvial geology. eds. J. K. Reed and M. J. Oard. Chino Valley, Arizona: Creation Research Society Books.
Underdown, S. 2008. A potential role for transmissible spongiform encephalopathies in Neanderthal extinction. Medical Hypotheses 71. no. 1:4–7.
Usshur, J. 1658. Rev. L. and M. Pierce. 2003. The annals of the world. Green Forest, Arkansas: Master Books.
Van Andel, T. H. and W. Davies (eds.). 2004. Neanderthal and modern humans in the European landscape of the last glaciation. McDonald Institute Monographs. Cambridge, England: McDonald Institute for Archaeological Research.
Van Bebber, M. and P. S. Taylor, 1994. Creation and time. Mesa, Arizona: Eden Productions.
Vieru, T. 2009. Neanderthal skull fragment dredged from North Sea. Retrieved October 30, 2009 from http://news.softpedia.com/news/Neanderthal-Skull-Fragment-Dredged-from-North-Sea-114301.shtml.
Von Fange, E. A. 1994. Noah to Abraham: the turbulent years. Published by author.
Walker, A. and P. Shipman. 1996. The wisdom of the bones: in search of human origins. New York: Alfred A. Knopf.
Weaver, T. D., C. C. Roseman, and C. B. Stringer. 2007. Were Neandertal and modern human cranial differences produced by natural selection or genetic drift? Journal of Human Evolution 53, no. 2:135–145.
Whitcomb, J. C. and H. M. Morris. 1961. The Genesis Flood: The biblical record and its scientific implications. Phillipsburg, New Jersey: P & R Publishing.
Wieland, C. 1994. New evidence: only people ever walked really upright. Technical Journal 8. no. 2:127–128.
Wieland, C. 2002. Skull wars: New Homo erectus skull in Ethiopia. Retrieved February 19, 2009 from http://www.icr.org/article/neanderthals-are-still-human/.
Wieland, C. 2008. A fusillade of shots in the Hobbit Wars. Journal of Creation 22, no. 3:25–27.
Wise, K. 2005. The Flores skeleton and human baraminology. Occasional Papers of the BSG 6:1–13.
Wise, K. 2008. Lucy was buried first: Babel helps explain the sequence of ape and human fossils. Answers 3. no. 2:66–68. Retrieved August 10, 2009 from: http://www.answersingenesis.org/articles/am/v3/n2/lucy-buried-first.
Wolpoff, M. H, B. Mannheim, A. Mann, J. Hawks, R. Caspari, K. Rosenberg, D. Frayer, G. Gill, and G. Clark. 2004. Why not the Neandertals? World Archaeology 36, no. 4: 527–546.
Wolpoff, M. H. and R. Caspari. 1996. Race and human evolution: A fatal attraction. New York: Simon and Schuster.
Wood, T. 2008. Four women, a boat, and lots of kids: A reevaluation of mitochondrial Eve. Answers 3, no. 2:70–74. Retrieved February 19, 2009 from http://www.answersingenesis.org/articles/am/v3/n2/four-women-boat-kids.
Woodmorappe, J. 2000. How different is the cranial-vault thickness of Homo erectus from modern man? Journal of Creation 14, no. 1:10–13.
Young, J. A. 2003. Septuagintal versus Masoretic chronology in Genesis 5 and 11. In Proceedings of the fifth international conference on creationism, ed. R. L. Ivey, Jr., pp. 417–430. Pittsburgh, Pennsylvania: Creation Science Fellowship.
Zilhão, J. 2006. Neandertals and moderns mixed, and it matters. Evolutionary Anthropology 15:183–195. doi:10.1002/evan.20110.














