
Yet Another ‘Missing Link’ Fails to Qualify
Originally published in Creation 15, no 3 (June 1993): 40-44.
A fossil truly ‘in-between’ the crucial fish and amphibian characters is not only hard to conceive, but has never been found.
After speaking at a recent public meeting on the campus of an Australian university, I was confronted by a palaeontologist from the local, large, publicly funded museum. He was irate at my assertion that the fossil record contained no evidence of major transitional forms or ‘missing links’ required by the evolutionary scenario, such as between fish and amphibians. He insisted that I was blatantly wrong and claimed that a ‘beautiful’ fossil record had been found in Greenland a few years ago which illustrated the fish-to-amphibians transition. This confrontation between us, and the palaeontologist’s claim, were subsequently featured in a write-up in a major Australian newspaper.1
This claim, of course, warranted full investigation. If verified, a series of fossils illustrating the transition between major types of organisms could prove to be a serious embarrassment to those who take God at His Word when He says He created separately the different types of creatures to reproduce only ‘after their kind’ (Genesis 1:21, 24, 25). Because God’s Word is true from the beginning (Psalm 119:160), I didn’t expect to find any proof for this claim. Besides, if this claim were true, then surely the evolutionist scientific community would be trumpeting the display of these fossils in every major museum and university, accompanied by bold headlines in major newspapers and popular scientific journals.
Of course, neither I nor others have seen such.
At the museum
I checked in the major Australian museum that employs this palaeontologist. Surely, if his claims were true, he would have featured this fossil series supposedly illustrating this major transition from fish to amphibians in his own museum.
Now in this museum there is a gallery on fossils and the geological record. Moreover, there is also a special exhibit entitled ‘Tracks Through Time—The Story of Human Evolution’, which features a section on fossils, including the transition from fish to amphibians. This special exhibition was launched in 1988 after being prepared under the direction of this palaeontologist. Five years later it is still on display in the museum, unchanged. Side-by-side are the rhipidistian fish Eusthenopteron and the amphibian Ichthyostega, the latter fossil having been found in the ‘Upper Devonian’ strata of East Greenland. These fossils (Figure 1) supposedly illustrate how fish evolved into amphibians. However, they fail to show how fins changed into legs. Missing is the claimed ‘beautiful’ fossil record found in Greenland which supposedly better illustrates this transition from fish to amphibians.
What the textbooks say
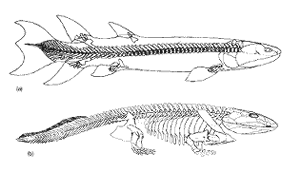
Figure 1. [Top to bottom]
(a) the rhipidistian fish Eusthenopteron. (b) The skeleton of the labyrinthodont amphibian Ichthyostega. Both originals were about one meter long. (After Carroll.3) Notice how completely are the fins and legs.
To confirm that this fish Eusthenopteron and amphibian Ichthyostega were not the transitional fossils (or ‘missing links’) claimed to have been found recently, I went to the textbooks on vertebrate palaeontology. Colbert,2 for example, in 1969 also used these fossils to illustrate the supposed transition from water to land. However, his accompanying diagram only illustrated similarities between the jaws and skulls of these fossils, and ignored the all-important claimed transition from fins to legs.
On the other hand, both Carroll3 in 1988 and Stanley4 in 1989 show drawings of the skeletal structures of the fins and legs respectively of these two fossils, making comparisons in order to illustrate how these fossils might represent the transition from the fish’s fin to the amphibian’s leg. Furthermore, in his text, Stanley says:
‘These fossils, many of which are assigned to the genus Ichthyostega, represent creatures that are strikingly intermediate in form between lobe-finned fishes and amphibians: The lobe fin itself is formed of an array of bones resembling that found in amphibians . . . . These features alone strongly suggest that amphibians were derived from lobe-finned fishes, but additional features make the derivation a certainty … Because of this intriguing combination of features, Ichthyostega, which was not discovered until the present century, represents what is commonly termed a “missing link”’.5
A pictorial diorama is then used by Stanley to reinforce this statement.
A palaeontologist’s admission
Stanley, who is on the staff of The Johns Hopkins University in Baltimore, USA, cannot be aware of the statement made earlier on this issue of ‘missing links’ by his colleague Dr Colin Patterson, Senior Palaeontologist at the British Museum (Natural History) in London. In 1978 that museum published a book on evolution by Patterson.6 Designed to be a popular book on the subject, it is still being sold in museums, even here in Australia. So it is still regarded as an authoritative presentation on evolution, including the fossil record. Yet, even though fossils are mentioned in a number of places in the book, nowhere does Patterson illustrate any ‘missing links’ between major types of organisms, such as between fish and amphibians.
In 1979 American Luther Sunderland read Patterson’s book and noticed this rather obvious lack of even a single photograph or drawing of a transitional fossil. So he wrote to Patterson asking why this omission, and in a letter dated 10 April 1979 Patterson replied in these words:
‘. . . I fully agree with your comments on the lack of direct illustration of evolutionary transitions in my book. If I knew of any, fossil or living, I would certainly have included them. . . . Gould and the American Museum people are hard to contradict when they say there are no transitional fossils . . . . You say that I should at least “show a photo of the fossil from which each type of organism was derived.” I will lay it on the line—there is not one such fossil for which one could make a watertight argument.’7
Greenland fossil finds
With this background I scanned the recent literature to see if any relevant new fossils had been found recently which might be the claimed ‘beautiful’ fossil record illustrating this fish to amphibians transition. Sure enough, in 1987 an expedition to Stensiö Berg in East Greenland by British scientists from Cambridge University and Danish scientists supported by the Greenland Geological Survey found very closely associated skulls of a new fossil, Acanthostega, at sites where fossil remains of Ichthyostega were also found.8
Ear bones and breathing
The first account of this new fossil material9 presented details of the skull and attempted to show that the middle-ear bone, while related to that in other tetrapods, had a functional part to play not only in hearing but also in breathing, which would make this bone similar to a bone in some fish that helps to pump in water, which is then expelled through the gill slits.10 It was also claimed that ‘The earliest tetrapods may have retained a fish-like breathing mechanism.’11 This naturally evoked scientific correspondence from other researchers,12,13 with a response from the Cambridge University palaeontologist.14
Fins and limbs
Next came a report from palaeontologist Clack and her colleague Coates at Cambridge University on the fossilized limb bones.15 They reported that the forelimb of Acanthostega had eight digits and the hindlimb of Ichthyostega had seven, quite unlike the common pattern of five digits on the feet (or hands) of many amphibians, reptiles, birds and mammals. They also described some resemblances of the forelimb skeleton of Acanthostega to the pectoral fin skeleton of Eusthenopteron, the similarities being viewed by the researchers in the context of the evolution of the tetrapod limb bones from the fin-bones of lobe-finned fishes (see Figure 2).
Such a statement is clearly based on the assumption of macroevolution, and not on observational evidence.
To account for this variation in digit numbers (from the general norm of five), Cooke16 suggested that conceivably the evolutionary process in the genetics of limb development in these ‘primitive’ amphibians was ‘not even well enough controlled to assure constancy between different individuals within single species’. He thus concluded that the common five-digit structure of tetrapod limb skeletons (the pentadactyl limb) must have become stabilized in a subsequent lineage, or lineages, to produce the common ancestors of today’s classes of tetrapods. Such a statement is clearly based on the assumption of macroevolution, and not on observational evidence of the bones in fins of fish changing into the limb bones of these amphibians and then other tetrapods.
A ‘missing link’?
In yet another paper Coates and Clack17 reported the discovery of what they regard as a fish-like gill (branchial) skeleton in Acanthostega, with grooves that they claimed are identical to those found in modern fishes. Thus they concluded that ‘Acanthostega seems to have retained fish-like internal gills . . . for use in aquatic respiration’.18 This they claimed ‘blurs the traditional distinction between tetrapods and fishes’ because it supposedly implies that the ‘earliest’ tetrapods were not fully land-dwelling (terrestrial). They further claimed that Acanthostega resembled a gill-breathing lungfish and that its legs with digits must have first evolved for use in water rather than for walking on land.19 They didn’t say it outright, but the implication is that they believe, as does the palaeontologist who confronted me with this example, that Acanthostega thus qualifies as a ‘missing link’ (transitional form).
No! A mosaic tetrapod
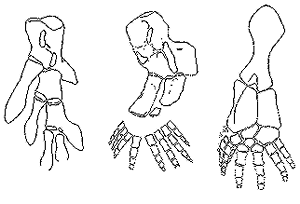
Figure 2. [left to right] (a) Pectoral fin skeleton of Eusthenopteron. (b) Restoration of the forelimb skeleton of Acanthostega. (c) Restoration of the hindlimb skeleton of Ichthyostega.
(All are dorsal view, anterior edges to the left, and drawn at a comparative scale only.) (After Coates and Clack.14) Notice that some of the Acanthostega limb bones are remotely similar to the Eusthenopteron fin bones, but the total limb is after the overall bone patter of fellow amphibian Ichthyostega. Notice also the varying digit numbers.
At about the same time, Clack and Coates made the following comment at an international conference:
‘Acanthostega gunnari is an Upper Devonian tetrapod which, like its better known contemporary Ichthyostega, displays a mosaic of fish- and tetrapod-like characters.’20
They also asked, rhetorically:
‘Was this animal secondarily aquatic, or do the fish-like characters indicate retention of the primitive condition? Were its tetrapod-like characters . . . evolved among more terrestrial tetrapods, or were they originally developed for life in shallow, swampy waters?’21
Clearly, in their minds, and the minds of their fellow evolutionary palaeontologists, this mosaic of fish- and tetrapod-like characters, and the presumed mode of life, make Acanthostega a ‘missing link’, even though they describe it as a tetrapod, that is, a four-legged animal. However, Acanthostega was a fully formed and fully functional four-legged amphibian, with four legs and not four fins, in some respects not unlike amphibians such as salamanders and newts.
Mosaics don’t count
Their description of Acanthostega as a ‘mosaic’ is significant. Acanthostega is not the first fossil to be called a mosaic, a creature that has characteristics common to two or more other types of creatures. For example, Australia’s platypus has milk glands and fur that classify it as a mammal, but it has a leathery egg, echo-location ability, a duckbill, webbed feet, poison spurs and other features that it shares in common with other animals, not only mammals. Like Acanthostega, Archaeopteryx has been regarded as an evolutionary intermediate (‘missing link’), but leading evolutionists Gould and Eldredge state that
‘Smooth intermediates . . . are almost impossible to construct, even in thought experiments; there is certainly no evidence for them in the fossil record (curious mosaics like Archaeopteryx do not count).’22
An amphibian nonetheless
Godfrey23 lists 41 characteristics that are unique to tetrapods. According to Ritchie,24 who has inspected the actual fossils, Acanthostega ‘fails the tetrapod test’ in eight out of these 41 characteristics, with two other characters not found in Acanthostega and another five not known from the fossil material. Thus Acanthostega still has 26 out of these 41 tetrapod characteristics. Ritchie also suggests that there are three other tetrapod characteristics present in Acanthostega not listed by Godfrey, so if these are included, Acanthostega has 29 out of 44 tetrapod characteristics. A 45th character which could be regarded as an unconventional tetrapod feature is the multi-digit, paddle-like limbs. On the other hand, Ritchie lists Acanthostega as having only eight potential ‘fish-like’ or ‘primitive’ characters.
However, the fact remains that Acanthostega has been classified as an amphibian (tetrapod) with a mosaic of tetrapod- and fish-like features. Nevertheless, leading evolutionists such as Gould and Eldredge regard mosaics as not qualifying as ‘missing links’. Interestingly, in his 1990 textbook Cowen25 doesn’t mention Acanthostega, even though reports on its claimed intermediate characteristics had appeared in the scientific literature from 1988 onwards.
Made up ‘stories’
So why do evolutionary palaeontologists and other scientists still persist in claiming that ‘missing links’ such as Acanthostega have been found, when some of their eminent colleagues have pronounced these fossils as failing to qualify? Again, Dr Colin Patterson’s comments are telling:
‘As a palaeontologist myself, I am much occupied with the philosophical problems of identifying ancestral forms in the fossil record. . . . It is easy enough to make up stories of how one form gave rise to another, and to find reasons why the stages should be favoured by natural selection. But such stories are not part of science, for there is no way of putting them to the test.’26
Several months later in an interview, after having been given two creation science books to read,27,28 Patterson was asked to elaborate, and in part of his response he said,
‘If you ask, “What is the evidence for continuity?” you would have to say, “There isn’t any in the fossils of animals and man. The connection between them is in the mind.”’29
In other words, fossils such as Acanthostega are regarded by some evolutionary palaeontologists as ‘missing links’ not because they are, but because they are believed to be. As Patterson says, it is ‘in the mind’, because ‘missing links’ are a philosophical necessity—to somehow provide ‘proof’ for their evolutionary faith.
Moreover,
‘The systematic status and biological affinity of a fossil organism is far more difficult to establish than in the case of a living form, and can never be established with any degree of certainty. To begin with, ninety-nine per cent of the biology of any organism resides in its soft anatomy, which is inaccessible in a fossil.’30
In any case,
‘. . . . anatomy and the fossil record cannot be relied upon for evolutionary lineages. Yet palaeontologists persist in doing just this.’31
Furthermore,
‘Everybody knows fossils are fickle; bones will sing any song you want to hear.’32
Conclusion
The fossil record has so far revealed many types of fish, some of which have bones in their fin lobes, serving a useful purpose as in the coelacanth (long believed to be an extinct ancestor of land animals, until it was found alive and well). The fossil record has also revealed many types of amphibians, including Ichthyostega and Acanthostega, in which the limb bones are firmly attached to the backbone and clearly designed for bearing the weight of the body in walking. Anything truly ‘in-between’ these crucial fish and amphibian characters is not only hard to conceive, but has never been found.
Footnotes
- Armitage, C., 1992. Beetle fires a blast in God versus evolution debate. The Sydney Morning Herald, Saturday, 22 August, p. 4.
- Colbert, E.H., 1969. Evolution of the Vertebrates, Second Edition, John Wiley and Sons, Inc., New York, pp. 76–78.
- Carroll, R.L., 1988. Verebrate Paleontology and Evolution, W.H. Freeman and Company, New York, pp. 156–166.
- Stanley, S.M., 1989. Earth and Life Through Time, Second Edition, W.H. Freeman and Company, New York, pp. 362–367.
- Stanley, Ref. 4, p. 364.
- Patterson, C., 1978. Evolution, British Museum (Natural History), London and University of Queensland Press, Brisbane.
- Sunderland, L.D., 1984. Darwin’s Enigma: Fossils and Other Problems, Master Book Publishers, San Diego (California), p. 89.
- Panchen, A.L., 1988. In search of earliest tetrapods. Nature, vol. 333, p. 704.
- Clack, J.A., 1989. Discovery of the earliest-known tetrapod stapes. Nature, vol. 342, pp. 425–427.
- Panchen, A.L., 1989. Ears and vertebrate evolution. Nature, vol. 342, pp. 342–343.
- Clack, Ref. 9, p. 427.
- Fritzsch, B., 1990. Evolution of tetrapod hearing. Nature, vol. 344, p. 116.
- Gottfried, M.D. and Foreman, B., 1990. Evolution of tetrapod hearing. Nature, vol. 344, p. 116.
- Clack, J.A., 1990. Early tetrapod hearing. Nature, vol. 344, p. 823.
- Coates, M.I. and Clack, J.A., 1990. Polydactyly in the earliest known tetrapod limbs. Nature, vol. 347, pp. 66–669.
- Cooke, J., 1990. Proper names for early fingers. Nature, vol. 347, pp. 14–15.
- Coates, M.I. and Clack, J.A., 1991. Fish-like gills and breathing in the earliest tetrapod. Nature, vol. 352, pp. 234–236.
- Coates and Clack, Ref. 17, p. 234.
- Coates and Clack, Ref. 17, p. 235.
- Clack, J.A. and Coates, M.I., 1991. Acanthostega—a fish out of water? In: D. Vezina and M. Arsenault (eds), 7th International Symposium on Studies of Early Vertebrates, Abstracts, p. 12.
- Clack and Coates, Ref. 20, p. 12.
- Gould, S.J. and Eldredge, N., 1977. Punctuated equilibria: the tempo and mode of evolution reconsidered. Paleobiology, vol. 3, p. 147.
- Godfrey, S.J., 1989. The postcranial skeletal anatomy of the Carboniferous tetrapod Greererpeton burkemorani Romer, 1969. Philosophical Transactions of the Royal Society of London, vol. B323, pp. 127–128.
- Ritchie, A., personal communication to W. Armstrong, October 1992.
- Cowen, R., 1990. History of Life, Blackwell Scientific Publications, Boston, pp. 145–158.
- Sunderland, Ref. 7, p. 89.
- Gish, D.T., 1973. Evolution: The Fossils Say No!, Creation-Life Publishers, San Diego (California).
- Wysong, R.L., 1976. The Creation-Evolution Controversy, Inquiry Press, Midland (Michigan).
- Sunderland, Ref. 7, p. 90.
- Denton, M., 1985. The fossil record. In: Evolution: A Theory in Crisis, Adler and Adler, Publishers, Inc., Bethesda (Maryland), p. 177.
- Lowenstein, J. and Zihlman, A., 1988. The invisible ape. New Scientist, vol. 120 (1641), p. 57.
- Shreeve, J., 1990. Argument over a woman. Discover, vol. 11(8), p. 58.

Support the creation/gospel message by donating or getting involved!

Answers in Genesis is an apologetics ministry, dedicated to helping Christians defend their faith and proclaim the good news of Jesus Christ.
- Customer Service 800.778.3390
- Available Monday–Friday | 9 AM–5 PM ET
- © 2026 Answers in Genesis