

Chapter 8
Differences Between Humans and Apes
How the fossil record, evolutionary assumptions, artistic license, and anatomical differences do NOT confirm common ancestry between humans and apes.

This chapter is from the book Searching for Adam, available in our online store.
Of all of the claims of biological evolution, perhaps none is more repugnant to conservative Christians than the bestial origin of man. Even when much of the Church had come to accommodate evolution in the late 1800s, there continued to be widespread resistance to the idea of the evolution of man from animals. For this reason, when Darwin wrote his Origin of Species in 1859, he chose not to include his views on human evolution. In a letter to the evolutionist Alfred Russel Wallace, Darwin explained that while he considered the evolution of man to be “the highest and most interesting problem for the naturalist” he would not discuss it in his book because the whole subject is “so surrounded by prejudices.”1 Darwin waited 12 years for the leaven of his Origin of Species to do its work before he published The Descent of Man (1871) in which he finally made public his own prejudices about human origins. Darwin confidently claimed that all the evidence pointed to man sharing common ancestry with the apes.
Darwin’s Lack of Fossil Evidence for Human Evolution
Darwin’s “evidence” in The Descent of Man for the common ancestry of man and apes consisted primarily of comparative anatomical, embryological, and behavioral arguments, rather than fossil evidence. Fossilized human skulls were found in Belgium in 1829 and Gibraltar in 1848 and later associated with the first recognized Neanderthal man fossils (discussed in the next chapter) found in Germany in 1856. But up until the time of Darwin’s death in 1882, no fossil evidence had been found for the supposed nonhuman or pre-human ancestors of man. Indeed, Darwin was puzzled by the lack of transitional fossils in general that might show how any two kinds of creatures could have evolved one into another. Darwin lamented, “Why, if species have descended from other species by fine gradations, do we not everywhere see innumerable transitional forms?”2 Darwin assumed that this lack of fossil evidence was a result of the incompleteness of the fossil record in his day and predicted that as more fossils were discovered, the “missing links” supporting his theory would eventually be found.
The Nature of the Fossil Record
Clearly, something about the fossilization process strongly favored the preservation of aquatic creatures, particularly bottom-dwelling immobile organisms with hard shells.
The fossil record favors the preservation of marine invertebrates. It is estimated that today over 250 million fossils have been found and cataloged, comprising about 250,000 species. Approximately 95% of all these fossils are marine invertebrates, and of the remaining 5%, most (95%) are algae and plants. Out of the less than 1% of all remaining fossils, 95% are other invertebrates (including insects). The vertebrates (animals with bones) comprise less than 0.25% of all fossils, and the vast majority of these are fish! Finally, primates (humans, apes, monkeys, and lemurs) comprise less than 0.001% of all vertebrate fossils.3 How then does one account for a fossil record where the vast majority of all invertebrates and vertebrates are aquatic? Clearly, something about the fossilization process strongly favored the preservation of aquatic creatures, particularly bottom-dwelling immobile organisms with hard shells.
The Process of Fossilization
The fossilization of plants and animals requires special conditions that generally include rapid burial in sediment, providing protection from predation and decay. These conditions are rarely met in a terrestrial environment. The vast majority of fossils were formed in layers of water-borne sediment. Where the sediment was sufficiently cementitious, the layers, with their imbedded fossils, become sedimentary rock. Stratified layers of fossil-bearing sedimentary rock cover about 75% of the earth’s land area, with many layers extending from continent to continent.
The global Flood of Noah’s day would have provided optimal conditions for depositing cementitious sediment and promoting fossilization on a global scale. While this explanation has long satisfied many Bible-believing Christians, evolutionists are loath to consider fossils being a result of a global flood. Nevertheless, even the vast majority of fossilized land-dwelling creatures were clearly in a water environment at the time of their burial in sediment. Evolutionists insist that these creatures were either the victims of local floods or that they wandered too close to an inland sea or lake and fell in. But animals caught in local floods or that fall into seas and lakes today rarely become fossils.
How Complete Is the Fossil Record of Land-Dwelling Creatures?
Creationists and evolutionists have very different expectations regarding the completeness of the fossil record. Evolutionists believe that a vast number of species evolved in the past 600 million years to produce the relatively fewer living species we have today. Using evolutionary assumptions, it has been estimated that less than 2% of all species that have existed on earth are preserved in the fossil record.4 Yet despite the assumed paucity of the fossil record, evolutionists expect to find fossils showing transitional stages in the progressive evolution of one kind of animal into a different kind.
Most creationists believe that essentially all of the fossils we see today were formed during and shortly after the global Flood that happened about 4,000 years ago. They believe that animals and plants have changed only to a limited degree within their created kinds since the Flood, though many have become extinct. Therefore, creationists would expect that fossils can be found that are recognizably similar to a majority of today’s living families, genera, and species. They would further expect that there would be no unambiguous fossil evidence of one kind of animal (such as a family or genus) progressively transforming into a different kind.
One way to estimate the relative completeness of the fossil record is to determine what percentages of the various kinds of animals living today have been found as fossils. If the fossil record were very incomplete, one would expect that relatively few representatives of today’s land-dwelling animals would be found as fossils. But this is not the case. Of the 43 orders of terrestrial vertebrates living today, one or more fossilized representatives have been found for 42 of the orders (97%). Of the 329 families of terrestrial vertebrates living today, fossilized representatives have been found for 261 families (79%).5
In a study focusing only on mammals, Bjorn Kurten reported that 88% of the living species of European mammals have been found in the fossil record of Europe, and 99% have been found in the worldwide fossil record.6 We may conclude that the fossil record appears to be remarkably rich and clearly sufficient to reveal if any creature has slowly evolved by intermediate stages into a distinctly different kind of creature.
Human and Primate Fossils Are Rare
Given the nature of the fossil record, it’s not surprising that human fossils are exceedingly rare.
Given the nature of the fossil record, it’s not surprising that human fossils are exceedingly rare. So rare indeed that most anthropologists teaching in our colleges and universities have never had opportunity to work with original primate fossils. Most are confined to examining published data and casts and reproductions of the actual fossils. Still, counting single teeth and bones, human fossils have been found representing more than 6,000 individuals.7 Apes and monkeys are also rare in the fossil record though they are more numerous than human fossils. It should be noted that evolutionists generally refer to apes in the fossil record as “apelike” creatures rather than apes. This avoids the embarrassment of saying apes evolved into apes.
Starting Assumptions about the Origin of Man
We often hear comments like “the fossils tell us,” but in and of themselves fossils don’t speak or tell us anything, including their age. Fossils exist in the present, and it is people who speak and give their interpretation of the past history of fossils based on their starting assumptions. The prevailing starting assumptions for understanding the fossil record are special creation and evolution.
Christian Assumptions about the Origin of Man
Most Bible-believing Christians in America believe that God created man and all other kinds of animals and plants in essentially their present form (generally represented by the family, not the species, taxonomic classification) by the power of His Word and Spirit. They observe and recognize the ongoing processes of extinction and limited variation within each kind, but point out that this has never been shown to produce fundamentally new kinds of creatures. Thus, they believe God created apes and God created men, but God did not create “ape-men” and apes didn’t change into man (with or without God’s providence). There are, however, a substantial number of professing Christians who believe God used evolution to “create.” But rarely do they specify what actual role God plays in evolution, or even if His existence is necessary for the evolution of the material cosmos.
Those Christians who attempt to accommodate evolution should reflect on the fact that professional evolutionists believe that religion itself is a product of evolution. Out of 14 billion years of purely materialistic cosmic evolution, religion is claimed to have evolved in the imagination of man’s ancestors only a few hundred thousand years ago. As the evolutionist Julian Huxley once put it, “Evolution is the whole of reality, a single process of self-transformation.”8 Theologians and Christian laymen should be aware that evolution is a jealous “god” that will have no other gods before it.
Evolutionist Assumptions about the Origin of Man
The foundational assumption of evolutionism is that evolution is a purely scientific and materialistic (naturalistic) explanation for the origin of everything that is real (i.e., the material universe). Divine intervention and intelligent design are anathema to nearly all evolutionary scientists. Nothing is considered to be above or outside of evolution, including the origin of man and his mental faculties. Even religion is considered to be a product of evolution. For example, an evolutionist from Humboldt University in Berlin observed chimps throwing rocks at trees for no apparent reason and concluded that this was a “worship ritual” telling us something about the evolution of religion. This was widely and enthusiastically reported in the popular media under the banner “Chimps believe in God!”9
Paleoanthropologists never ask the question, “Did man evolve from apelike creatures?”
Evolutionists assume that amoeba-to-man evolution is an absolute fact, though they concede that the details regarding the actual mechanism of biological evolution and what creature evolved into what are theoretical or even speculative. In the case of human evolution, it is considered an unassailable fact that humans have evolved from nonhuman ancestors. Thus, paleoanthropologists never ask the question, “Did man evolve from apelike creatures?” Indeed, such a question would likely be career-ending. Paleoanthropologists need only concern themselves with which of the apelike creatures that have been found in the fossil record are our ancestors. They are certain that at least some of these fossil apes must be our ancestors or else we wouldn’t have any apelike ancestors, and that’s unthinkable to an evolutionist.
Another primary assumption of evolutionism is that the degree of anatomical, functional, and genetic similarity between two creatures is considered evidence of their degree of evolutionary relatedness. For example, humans are obviously more similar to apes than they are to fish, so our presumed evolutionary relationship to apes is considered to be much closer than it is to fish. But even fish are considered to be our distant relatives because we both have vertebrae and other similarities common to vertebrates.
But there are countless examples of striking structural and functional similarities between unrelated organisms that make no sense in terms of evolutionary relatedness. For example,
- Opossums and primates have an opposable thumb.
- Australian koalas have fingerprints almost indistinguishable from humans.
- Bats, whales, and shrews have similar sonar-like echolocation.
- Among vertebrates, only mammals and certain salamanders and fish have non-nucleated red blood cells.
- There are many strikingly similar pairs of marsupial and placental mammals, yet evolutionists believe the two separated 160 million years ago.
- A single cell dinoflagellate (protozoan) has been found with a vertebrate-like camera eye consisting essentially of a cornea, lens, and retina derived from subcellular organelles including mitochondria and plastids.10
All of these similarities are “explained” by evolutionists with a rescuing hypothesis called “convergent evolution.” Convergent evolution is when two distantly related or unrelated creatures are claimed to have independently evolved by chance the same trait or traits. It would seem that evolutionists consider anything possible, even probable, except intelligent design of biological systems by our Creator.
Comparative Anatomy of Apes and Humans
Most of the fossil evidence for human evolution is based on the comparative anatomy of primates and humans. This requires a knowledge of the anatomy of both living and fossil primates. The hard tissues (teeth and bones) are particularly important since primate fossils are generally partially preserved, disarticulated skeletons that rarely show evidence of soft tissues like the organs of the body.
The layman considering the fossil evidence for human evolution will be overwhelmed with unfamiliar anatomical terms. But there are some basic differences in the hard tissue anatomy of apes and humans that can be easily understood and that clearly distinguish apes from humans. This knowledge can be insightful when one visits a human evolution display at a natural history museum or examines illustrations and descriptions of presumed ape-men in the popular media. Indeed, the whole purpose of this chapter is to help the non-specialist critically evaluate the current and future fossil evidence for the presumed “missing links” of human evolution.
Skulls - Differences Between Humans and Apes
Small Brain vs. Big Brain
Skulls are perhaps the most interesting primate fossils because they house the brain and give us an opportunity, with the help of imaginative artists, to look our presumed ancestors in the face. The normal human skull is easily distinguished from all living apes, though there are of course similarities.
The cranial capacity of the skull is large in humans because of their relatively large brain compared to that of apes. The average brain size of an adult human is about 1,400 cm3, but can vary among individuals over a nearly threefold range from about 700 cm3 to 2000 cm3. It should be noted that the size range of the normal human brain does not correlate with intelligence. Rather, brain size generally correlates with body size. As a result, adult human males have a brain volume that averages about 100 cm3 larger than the brain of adult human females, reflecting the relative difference in their average body size.
Adult apes have brains that are generally about one-third the size of normal adult human brains. As in humans, the brain size of apes varies with body size and is generally in the range of 300–500 cm3. But the profound difference in intelligence between apes and humans is not a simple matter of the human brain being three times larger. The difference in the complexity of the interconnections between individual brain cells, for example, is far more important in cognitive ability than mere brain size. This important difference is not perceivable in fossils.
Position of the Foramen Magnum
The position of the foramen magnum of the skull is often used as an indicator of bipedality (habitually walking on two legs), since it usually indicates the orientation of the head to the trunk of the body. The foramen magnum is a large oval opening in the occipital bone on the base of the skull where the spinal cord exits the cranial cavity. This opening is generally located farther forward in upright walking bipeds and farther back on the skull in quadrupeds. This difference is obvious when comparing non-primate quadrupeds like a dog to a human. But the situation is more complicated in primates.11 There are two common methods for quantifying the relative position of the foramen magnum in primates: the basion-biporion distance and the basion-bicarotid chord distance. These measurements can give conflicting results.12 The basion-bicarotid method, for example, distinguished between apes and humans, but the basion-biporion method is less successful. In the case of chimpanzees, the basion-bicarotid method shows a sexual difference in the relative position of the foramen magnum being more anterior in males than females.
Sloped vs. Vertical Face
Perhaps the easiest way to distinguish an ape skull from a human skull is to examine it from a side view (Figures 1 and 2). From this perspective, the face of the human is nearly vertical while that of the ape slopes forward from below the eye sockets to the tip of its upper jaw. The easiest way to evaluate the slope of the face in a skull is to compare the angle of the face to the zygomatic arch (cheek bone). The human face is nearly perpendicular to the zygomatic arch while the ape face is sloped at an obviously oblique angle. By this criterion alone, one can easily distinguish the skulls of living apes from humans.
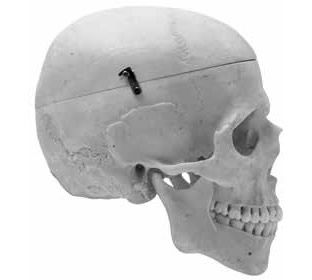
Figure 1: Human
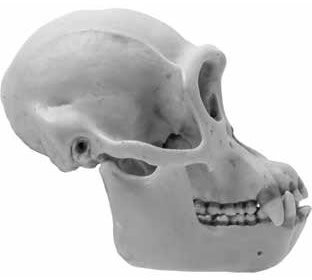
Figure 2: Chimp
Flat vs. Curved Forehead
The skull above the base of the orbits (eye sockets) is flat in apes and curved in humans. From a side view of the skull, the bony socket of the ape’s orbit is obscured by its broad, flat, upper face. Humans, on the other hand, have a horizontally curved upper face and forehead, clearly revealing the orbit of the eye from a side view (figures 1 and 2). Also, when viewed from the front, the orbit of the human eye is slightly wider than it is tall, while the orbit of apes is usually slightly taller than wide, making them look goggle-eyed.
Flat vs. Protruding Nasal Bones
The nasal bones are a pair of bones meeting in the midline covering the upper part of the nasal cavity. Human nasal bones are distinctive in that they protrude from the face (the nose bones that eyeglasses rest on) [Figures 3 and 4]. Apes, by comparison, have flat nasal bones. As a result, the slight protrusion of the ape’s nose is made up entirely of soft tissue and cannot be fractured like the human nose. Another difference is that humans have a protruding nasal spine, which anchors the nasal septum, while apes lack such a structure. The human nasal spine can be felt by placing your finger across your upper lip under your nose and then attempting to raise the finger.
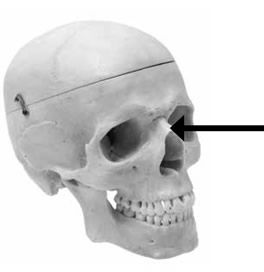
Figure 3: Human nasal bone (see arrow)
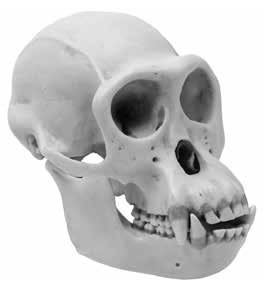
Figure 4: Chimpanzee (with flattened nasal bone)
Jaws and Teeth
Because of their relative hardness, teeth and jaw fragments are the most frequently found primate fossils. Tooth enamel is the hardest biological substance known. Essentially, all of the living cells and organic tissue that produce the enamel in the developing tooth are replaced by mineral crystals of hydroxyapatite. One might say that the enamel of our teeth is one part of our body that is fossilized while we are still alive. Thus, much of the evidence for the ape ancestry of man is based on similarities of teeth and jaws.
In contrast to man, apes tend to have incisor and canine teeth that are relatively larger than their molars. Ape teeth usually have thin enamel, while humans generally have thicker enamel. Finally, the jaws tend to be more U-shaped in apes and more parabolic in man, though there can be considerable variation among individuals of a species.
The problem in declaring a fossil ape to be a human ancestor (i.e., a hominin) on the basis of certain human-like features of the teeth is that some living apes have similar features and no living ape is considered to be an ancestor of man. Some species of modern baboons, for example, have relatively small canines and incisors. While most apes do have thin enamel, some apes such as the orangutans have relatively thick enamel. Teeth are likely to tell us more about an animal’s diet and feeding habits than its supposed evolution. Nonetheless, relatively thick enamel is a commonly sighted criterion for declaring an ape fossil to be a hominin.
Pelvis and Legs - Differences Between Humans and Apes
Humans are the only living primates that habitually walk on two legs.
Humans are the only living primates that habitually walk on two legs. Thus, the most eagerly sought-after evidence in fossil hominins is any anatomical feature that might suggest bipedality (the ability to walk on two legs). Any hint, real or imagined, of bipedal locomotion in fossil apes is considered by evolutionists to be compelling evidence for their status as human ancestors. Nonhuman primates generally walk in an essentially quadrupedal fashion, using both their feet and hands. Only the bonobo (pygmy chimpanzee) is capable of extended (25% of the time) bipedal locomotion. While nonhuman primates do occasionally walk briefly on two legs, their manner of walking is entirely different from that of humans. The distinctive human gait requires the complex integration of many skeletal and muscular features in our hips, legs, and feet. Thus, evolutionists closely examine the hipbones (pelvis), thighbones (femur), leg bones (tibia and fibula), and foot bones of fossil apes in an effort to detect any anatomical features that might even remotely suggest bipedality.
Pelvis
The pelvis (hip bones) plays a critically important role in walking, and the distinctive human gait requires a pelvis that is anatomically and functionally different from that of the apes. Indeed, one has only to examine the pelvis to determine if any hominin was likely to have had the ability to walk like a human.
The part of the hip bone that we can feel just under our belt is called the ilium or the iliac blade. Viewed from above, human ilia are curved forward like the handlebars of a tricycle. The iliac blades of the ape, in contrast, project straight out to the side like the handlebars of a scooter (figures 5 and 6). This is important because the shape of the iliac blades of the hipbones determines the function of an important pair of muscles called the gluteus medius and clearly reveals if a creature is capable of human-like bipedal walking.
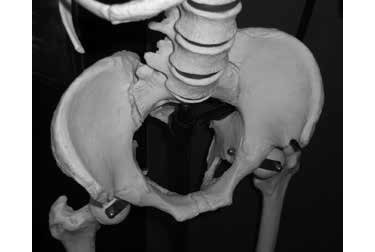
Figure 5: Human iliac blades
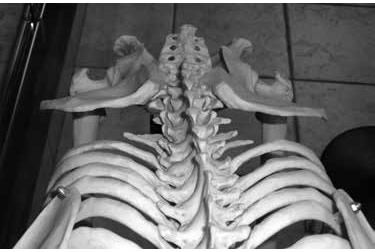
Figure 6: Chimpanzee iliac blades
In both apes and humans, the gluteus medius has its origin on the superficial surface of the iliac blade and inserts into the greater trochanter of the femur. Because of the anteriorly curved iliac blade of humans, the gluteus medius muscle runs down the side of the hip (like a holster) to where it inserts on the greater trochanter of the femur. In the case of the ape with its straight iliac blade, the gluteus medius runs dorsally down the back from its origin in the iliac blade to its insertion on the greater trochanter of the femur. This results in two distinctly different actions of the human vs. the ape gluteus medius. The human gluteus medius serves as an abductor of the leg (swings the leg out to the side). The ape gluteus medius serves as an extensor of the hip, which brings the upper body of the forward-leaning ape to a more vertical position.
In the human, the gluteus medius stabilizes the hip when walking. This is important because the articulation of the head of the femur with the pelvis is a ball-socket type joint with a wide range of mobility. When we lift our left foot off the ground to make a step, our hip would fall on the left side were it not for the contraction of the gluteus medius on the side of our planted foot.
Because of the shape of the pelvis and orientation of the gluteus medius muscles, humans are able to walk while keeping the left and right hips essentially parallel to the ground. When apes walk on two legs, however, they must swing their upper body from side to side in the familiar “ape walk” fashion, to keep their weight over the planted foot. The reason for this difference in gait is largely a matter of pelvic anatomy resulting in a difference in the orientation of the gluteus medius muscles.
Carrying Angle of the Knee
Evolutionists are particularly interested in the angle at which the femur and the tibia meet at the knee (called the valgus or carrying angle). Humans are able to keep their weight over their feet while walking because our femurs converge toward the knees, forming a carrying angle of approximately nine degrees with the tibia, making us sort of knock-kneed (figure 7). In contrast, chimps and gorillas have widely separated straight legs with a carrying angle of essentially zero degrees.
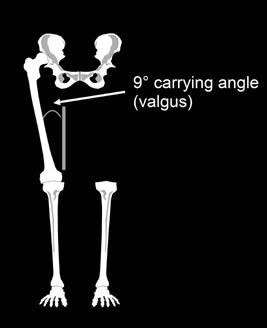
Figure 7: Human carrying angle
Evolutionists assume that fossil apes with a high (human-like) carrying angle were bipedal and thus evolving into man. For example, certain australopithecines (an apelike creature) are considered to have walked like us and thus to be our ancestors largely because they had a high carrying angle. But high carrying angles are not confined to humans. They are also found in some modern apes and monkeys that walk gracefully on tree limbs and only clumsily on the ground.
Living apes and monkeys with a high carrying angle (values comparable to man) include apes such as the orangutan and monkeys such as the spider monkey—both adept tree climbers and capable of only an apelike bipedal gait on the ground. The high carrying angle in these creatures allows them to keep their feet close together when walking on limbs of trees. The point is that there are living tree-dwelling apes and monkeys with some of the same anatomical features that evolutionists consider to be definitive evidence for bipedality, yet none of these animals walks like man and no one suggests they are our ancestors or descendants.
Locking Knee
Another distinctive difference between the legs of humans and apes is that the human knee locks when the leg is straightened and unlocks when the knee is bent. The lock is achieved by the tibia rotating about five degrees outward in relationship to the femur when the leg is straightened at the knee. This explains why our footprints point outward at the toes. Apes lack this locking mechanism and must walk with slightly bent knees. Humans have only to walk a few minutes with slightly bent knees to appreciate the value of locking knees. It requires considerably more energy to support walking on two legs when the legs are even slightly bent at the knee.
The locking knee is also of value when standing for a long period of time. When standing at attention, for example, our toes point forward but our femurs rotate inward relative to the tibia and the lock is set. Thus when a prankster clips a standing person behind the knee, it can cause them to fall before they can reset the lock.
Foot Bones
The human foot is unique compared to any other primate because it is designed specifically for walking, while the feet of other primates are used for grasping as well as walking.
The human foot is unique compared to any other primate because it is designed specifically for walking, while the feet of other primates are used for grasping as well as walking. The big toe of the human foot is inline with the foot, not jutting out to the side like an ape foot. Human toe bones are relatively straight rather than curved and grasping like ape toes. The human big toe points forward and is in alignment with the other toes. The big toe of the apes, by contrast, angles inward from the rest of its toes and is a grasping toe used almost like a thumb in climbing trees.
Another distinctive feature of the human foot is its complex arches. Our foot has three distinct arches, the longitudinal medial and longitudinal lateral arches running from the heel to the base of the toes on each side of the foot, and one transverse arch running across the mid foot from medial to lateral sides. These arches are made up of bones, ligaments, and tendons, and are essential for bearing weight and forward propulsion in bipedal locomotion. To serve these functions the foot must be highly stable yet flexible. All living apes by comparison are flat-footed and lack the longitudinal and transverse arches.
When a human walks, the heel of the foot hits the ground first, then the weight distribution spreads forward from the heel along the outer margin of the foot up to the base of the little toe. From the little toe the weight-bearing spreads inward across the base of the toes and finally pushes off with the big toe. No ape has a foot or push-off like that of a human and thus no ape is capable of walking with the distinctive human stride or capable of making human-like footprints. As a result, a series of fossilized footprints can provide compelling evidence for both foot anatomy and stride. But as we will see, when evolutionists encounter footprints that appear to be human in both anatomy and stride, but are considered too old to be human, they attribute them to apelike hominins, not humans.
Hand Bones - Differences Between Humans and Apes
The human hand is easily distinguished from that of nonhuman primates. While the human hand is designed to serve exclusively for grasping and manipulating, the hand of other primates is designed to serve both as a hand and a foot. Nonhuman primates use their hands to either fist walk (bonobos and orangutans) or knuckle walk (chimpanzees and gorillas).
The thumb is perhaps the most distinctive feature of the human hand. Our thumb is longer than that of any other primate, extending well beyond the knuckle of our forefinger. All other primates have a shorter thumb that barely reaches the knuckles. Most primates have opposable thumbs, but none have the range of motion of the human thumb. The human thumb is more muscular and mobile, and unlike apes, can touch all four of our fingers.
Human finger bones are relatively shorter, straighter, and flatter than the fingers of nonhuman primates, which have longer and more curved fingers. Finger curvature is very pronounced in knuckle-walkers and in the suspensory adapted hand of tree climbers. In addition, the wrist (carpal) bones of knuckle-walkers can move into a configuration that locks the wrist.
Scapula (Shoulder Blade) - Differences Between Humans and Apes
In all primates (and limbed vertebrates in general), the only bony attachment of the arm to the rest of the skeleton is by way of the scapula (shoulder blade) and clavicle (collar bone). The humerus (upper arm bone) articulates with the scapula at a shallow ball-socket joint called the glenoid cavity. The scapula articulates with the clavicle, which in turn articulates with the manubrium of the sternum (upper end of the breast bone). This arrangement gives a much greater range of motion to the upper limbs than is possible with the deep ball-socket joint of the lower limbs.
In the case of humans, the joint between the humerus and the scapula only allows us to raise our arm to a position approximately parallel to the ground. Any further elevation of the arm requires the scapula to be rotated by means of muscles on our shoulder and back. Thus it requires more effort to use our arms over our head than to use them below shoulder level.
Arboreal apes and monkeys have suspensory adapted forelimbs that are often used in a highly elevated position when grasping limbs of trees. This is facilitated in primates by an up-tilted glenoid cavity that permits the forelimbs to be elevated over the head with little or no rotation of the scapula. Thus the anatomy of the scapula can reveal if a primate is a tree-dweller.
Other Variables That Must Be Considered
Sexual Dimorphism
The anatomical differences between the human male and female skeleton are subtle though apparent to the trained eye, but the differences between male and female ape skeletons are usually much more obvious. These skeletal differences are called sexual dimorphism. Sex-related differences in nonhuman primate anatomy are particularly common and particularly evident in the skull.
If one compares a male gorilla skull with that of a female gorilla, the male skull is much bigger (after taking account of differences in body size) and more robust in appearance. Another major difference is that the male gorilla skull has a much larger sagittal crest than the female does. The sagittal crest is a bony ridge or plate that runs lengthwise along the midline on the crest of the skull. This bony ridge serves to anchor jaw (chewing) muscles called the temporalis muscles. Humans, by contrast, have nothing resembling a sagittal crest because their much smaller and thinner temporalis muscle attaches to the side of the skull (temporal region). A noticeable sagittal crest is found on the males of the more robust apes such as the gorilla and orangutan but occurs only rarely among the males of smaller apes such as the chimpanzee.
Age Differences
Bones are very dynamic structures that change in shape and density both with age and use. The primate skeleton in particular shows substantial developmental change with age. The skull of juvenile primates, for example, is relatively large for its body size compared to adults, due to the relatively rapid early development of the nervous system. Thus a juvenile ape might appear to have a relatively large brain for its body size and be interpreted as human-like, if age and development are ignored.
Pathology
Pathological conditions can profoundly change the anatomy of bones. Failure to consider pathological change can lead to a gross misinterpretation of fossil primates. For example, an early Neanderthal fossil known as La Chapelleaux-Saints showed obvious osteo-degenerative disease including loss of teeth, advanced resorption of mandibular bone, and advanced arthritis. A reconstruction of this specimen by Marcellin Boule in 1911 failed to take pathology into account, which led to portraying Neanderthals as having a stooped posture, a thrust-forward skull, bent knees, and divergent big toe. As a result of this misinterpretation and evolutionary bias, Neanderthals were not considered to be direct ancestors of humans until the fossil was properly re-examined by Straus and Cave in 1957 and shown to be human.
Fragmentation of Fossils
Primate fossils are often highly fragmented and rarely articulated. The famous KNM-ER 1470 skull, for example, was assembled from more than 150 fragments. Accurate assembly requires a thorough knowledge of primate anatomy, a good eye, and a freedom from observer bias. The skull of 1470 is classified as Homo habilis (or Homo rudolfensis), but some evolutionists believe it is an australopithecine. The confusion arises in part from the fact that the cranial capacity of skull 1470 is quite large (800 cm3) for the size of the skull, but the skull has been reconstructed with a sloping face giving it an apelike appearance. However, in this specimen the only attachment of the face below the orbits of the eyes to the rest of the skull is a slender bone fragment that allows the lower face to be reconstructed as either sloped like an ape or vertical like a human.
Three Ways to Make an Ape Man
Both apes and humans are found in the fossil record, but is there really compelling evidence for fossilized ape-men?
Both apes and humans are found in the fossil record, but is there really compelling evidence for fossilized ape-men? Evolutionists use three ways to construct ape-men from ape and human fossils:
- Combine ape and human fossils declaring them to be one individual
- Upgrade fossil apes to ape-men
- Downgrade fossil humans to ape-men
Combining Fossil Bones of Apes and Humans to Make Ape-men
One way to make an “ape-man” is to combine fossilized ape bones with fossilized human bones and then declare the mixture to represent one individual. This could be either intentional deception or an unintentional mistake.
The most famous example of an ape-man proven to be a combination of ape and human bones is “Piltdown Man.” In 1912, it was announced at the Geological Society of London that Charles Dawson, a medical doctor and an amateur paleontologist, had discovered a mandible (lower jaw) and part of a skull in a gravel pit near Piltdown, England. A canine tooth was found later and added to the collection. The jawbone was apelike, but the teeth were described as showing wear similar to the human pattern. The skull fragments, on the other hand, were very human-like, indicating a very large brain. These three specimens were combined to form what was also called “Dawn man,” and was confidently dated to be 500,000–1,000,000 years old, which “afford us a link with our remote ancestor, the apes.”13 The fossils went into the Natural History Museum (NHM) in London, and only plaster casts of the evidence were shown to scientists or the public.
Finally, in 1953, after scientists outside the NHM examined the actual fossils, the whole thing was exposed as an elaborate hoax perpetrated by someone who knew what evolutionists were looking for. The skull was indeed human (dated to be about 500 years old), while the jaw was that of a modern female orangutan whose teeth had been obviously filed to crudely resemble the human size and wear pattern, and the canine tooth came from a chimpanzee. The long chimp canine tooth was filed down so far that it exposed the pulp chamber, which was then filled in by the hoaxer in an apparent effort to hide the mischief. It would seem that any competent scientist examining this tooth would have concluded that it was either a hoax or the world’s first root canal. But the deliberate hoax apparently involved some of the leading scientists in Britain.14 The success of this hoax for over 50 years in spite of 500 research papers written on the find15 and the apparently careful scrutiny of some of the best authorities in the world led the prominent evolutionist Lord Solly Zuckerman to declare, “Students of fossil primates have not been distinguished for caution when working within the logical constraints of their subject. The record is so astonishing that it is legitimate to ask whether much science is yet to be found in this field at all. The story of the Piltdown Man hoax provides a pretty good answer.”16
The fossil bones of apes and humans are rarely found in an articulated condition. Fossilized primate bones are often widely scattered. Where several bones are found in one location, it can be difficult to determine if they are parts of one individual creature or several. Fossil bones believed to be from one individual may be widely scattered and discovered over a period of months and years.
Upgrade Fossil Apes to Ape-men
Another way to make an “ape-man” is to find something about the apes in the fossil record that can be used in an effort to promote them to a more human-like status. Since even living apes and humans share many skeletal similarities, it is quite easy to make a list of similarities between fossil apes and humans. But these similarities do not prove an evolutionary relationship. There are also distinctive features of the human skeleton that make it quite easy to distinguish today’s humans from today’s apes. The two-part question is simply this: are fossil apelike creatures really significantly different from modern apes, and are they really more similar to humans?
Of the four genera of hominins that evolutionists believe to be ancestral to humans, three are clearly very apelike. The ardipithecines, australopithecines, and paranthropithecines are apes and would doubtless be recognized as such if they were living today. Not only is their anatomy very apelike, but their fossils are never directly associated with human artifacts such as tools, art, hunting weapons, ritual burials, lodgings, or use of fire. The jump from these apelike creatures to members of the genus Homo is dramatic and consistent with the fact that God created apes and men but not “ape-men.”
Downgrade Fossil Humans to Ape-men
There are clearly fossil humans in the fossil record, though they are rare. But if there is to be evidence for evolution of humans from nonhumans, fossils of “primitive” humans must be found with differences in anatomy that are apelike and clearly fall outside the normal range of human variability.
When these human fossils are considered to be too old to be human, they become ape-men regardless of their anatomy.
When these human fossils are considered to be too old to be human, they become ape-men regardless of their anatomy. Thus, the obviously human-like footprints found by Mary Leakey at the Laetoli site in Tanzania are considered to be made by the very apelike Australopithecus afarensis because they have been “dated” at 3.66 million years old and thus are too old to be human, in spite of the fact that they look so human in both anatomy and stride (see below).
The Role of the Artist in Promoting Human Evolution
Several things should be noted about the artistic reconstructions of ape-men that we find in museum displays, textbooks, and news articles.
The “March of Progress”
Nowhere is the key role of the artist in promoting belief in evolution more evident than in an illustration published in 1965 in the Time-Life Book Early Man, featuring a foldout illustration of 13 presumed ancestors of man over 25 million years of evolution.17 As one progresses from left to right, the “ancestors” stand a little taller and more erect and assume a progressively more human-like posture and gait (Figure 8). Such a linear and progressive ancestor-descendant parade was known to be untrue even at the time it was drawn, but has nonetheless become an enduring icon of evolution known as the “March of Progress.”

Figure 8: “The Road to Homo sapiens”
Many consider the “March of Progress” to be the most famous and influential image in the history of scientific illustration. It has been copied in a shorter form on countless T-shirts, buttons, book covers, record albums, commercial advertisements, programs for scientific meetings, and is even used as the emblem of the Leakey Foundation. As a result of this wide exposure, many laymen base their belief in the evolution of man more on this easy-to-understand image of linear “progress” than on any actual scientific evidence. But evolutionary anthropologist Bernard Wood informs us,
There is a popular image of human evolution that you’ll find all over the place. . . . On the left of the picture there’s an ape . . . On the right, a man . . . Between the two is a succession of figures that become ever more like humans. . . . Our progress from ape to human looks so smooth, so tidy. It’s such a beguiling image that even the experts are loath to let it go. But it is an illusion.18
The Whites of Their Eyes
Artistic renderings of fleshed-out fossils of presumed ancestors of man often show the face of a very apelike creature with very human-like eyes. This is accomplished by merely putting a white sclera in the eyes (the whites of the eye). All apes and monkeys have a brown sclera that is nearly as dark as their brown iris, while all humans with normal, healthy eyes have an essentially white sclera. Obviously, the fossil record for scleral color is nonexistent, so this is simply artistic license used to influence the viewer.
Artistic Extrapolation
In his book Life on the Mississippi, Mark Twain wryly commented, “There is something fascinating about science. One gets such wholesale returns of conjecture out of such a trifling investment of fact.” Nowhere is this more evident than in the field of evolution. Where data are missing, evolutionists often employ artists to fill in all the desired missing evidence. Artistic imagination has been used to illustrate entire “ape-men” from nothing more than a single tooth. In 1922, Henry Fairfield Osborne, director of the American Museum of Natural History, reported on a molar tooth from the Pliocene strata of Nebraska that he claimed had characteristics of both man and ape. The tooth was given the scientific name Hesperopithecus but became commonly known as “Nebraska man.” The London Illustrated News (LIN) even showed a fleshed-out, double-page illustration of Nebraska man and his wife in their natural habitat!
“Nebraska man” along with “Piltdown man” (now known to be a hoax) were the prevailing evidence for human evolution during the time of the famous Scopes “monkey trial” in 1925. It is said that even many theologians of the time accepted the evolutionary claim for the bestial origin of man on the evidence of these imaginary ape-men. In 1927, parts of the skeleton of Hesperopithecus were discovered together with more teeth, and Nebraska man was found to actually be an extinct peccary (wild pig)! Making an ape-man out of a pig tooth is an example of the extreme observer bias often found in paleoanthropology. One might sum up this bias as, “I wouldn’t have seen it if I hadn’t believed it.” The reinterpretation of the fossil evidence was announced in a technical journal, the popular picture from the LIN was quickly removed from literature, and the general public was never the wiser.19
The Fossil Hominins (Man and His Apelike “Ancestors”)
The reader should now have sufficient background to critically examine specific examples of the current crop of presumed apelike ancestors of humans.
The reader should now have sufficient background to critically examine specific examples of the current crop of presumed apelike ancestors of humans. There will no doubt be many more proffered examples of “missing links” between apes and humans in the future. The popular media will be eager to present each new “ape-man” to you with the assurance that this is finally the true missing link that all evolutionists have been waiting for. But you must learn to be critical, think for yourself, and dare to question scientific (or theological) authority. The hominin fossils discussed in this chapter follow the organization and nomenclature of an exhibit from the Smithsonian National Museum of Natural History (entitled “What does it mean to be human?”) that is traveling to public libraries across America.20
The Classification of Hominins
The taxonomic classification of living plants and animals has become chaotic in recent years. In the past, classification was largely based on anatomical similarities. But now embryological, genetic, and molecular similarities, as well as presumed evolutionary relationships are considered in classification. But these different approaches frequently lead to very different classification schemes. When such differences arise, assumptions about evolutionary relatedness generally trump everything else.
The classification of man’s presumed fossil ancestors is chaotic at best. One problem is that the nomenclature is constantly changing. Currently, the term “hominid” refers to modern humans as well as all modern and extinct Great Apes and their immediate ancestors. A newer term “hominin” refers to modern humans and all of our presumed ancestors, which include the genera Homo, Paranthropus, Australopithecus, and Ardipithecus. Note that humans are the only living hominins; all other hominins are speculations from the fossil evidence. This classification is obviously based on evolutionary starting assumptions.
The Common Ancestor of Apes and Humans
While evolutionists are certain that modern apes (like the chimpanzee) and modern humans shared a common ancestor six to seven million years ago, this ancestor has never been specifically identified. Several fossilized apelike creatures have been proposed as being ancestral to humans, but rarely are fossil apelike creatures identified as being only ancestral to modern apes. It would appear that when paleoanthropologists are lucky enough to find a rare fossil apelike creature, it will not be wasted on simply being an ancestor of apes when with a little more effort it can be declared to be an ancestor of humans. There is obviously more fame and fortune to be had in finding a supposed ape ancestor of humans than an ape ancestor of apes.
In a 2015 study published in the Proceedings of the National Academy of Sciences,21 evolutionists suggest the common ancestor of apes and humans looked a lot like a chimpanzee or a gorilla—they’re just not sure which. Is there any other field of empirical science where something this tenuous and trivial would be accepted for publication in the prestigious PNAS?
Ardipithecus Group
The Ardipithecus group is considered by evolutionists to be the very earliest apelike human ancestors, which evolved in Africa four to six million years ago. The genus name Ardipithecus is derived from an Afar Ethiopian word, ardi, which means “ground” or “floor,” and Greek word pithekos, which means “ape.” Thus the name Ardipithecus or “ground ape” reflects the evolutionists’ belief that these were the first apelike creatures to begin to habitually walk upright on the ground. All the adult members of this group are very apelike and are about the size of chimpanzees.
Ardipithecus ramidus
The first fossils named Ardipithecus were found in 1994 by paleoanthropologist Tim White in the Middle Awash region of Ethiopia. Initially, the fossil was attributed to the Australopithecus genus, but was later declared to be a new genus, Ardipithecus. The species name ramidus is derived from “ramid,” which in the Afar language of Ethiopia means “root,” referring to the belief that this ape is at the very root of humanity. White and his co-workers have found about 100 partial fossil specimens that are attributed to Ar. ramidus.
Evolutionists claim that Ar. ramidus is 4.4 million years old, making it over a million years older than the famous australopithecine fossil called “Lucy,” which had long been believed to be the oldest hominin to walk on two legs. But the evidence for anything resembling human-like bipedality in “Ardi” is hardly compelling. First, the foot bones of this creature reveal a very apelike, divergent, grasping big toe and rigid foot. The pelvis was reconstructed from crushed fragments and, according to some evolutionists, retained considerable arboreal capabilities and is at most only suggestive of bipedalism.
Ardipithecus kadabba
The fossil known as Ar. kadabba was found in the Middle Awash region of Ethiopia by paleoanthropologist Yohannes Haile-Selassie in 1997. It was originally considered to be a subspecies of Ar. ramidus, but was later declared to be a separate species on the basis of perceived differences in its teeth. The first specimen found was a lower jaw, but eventually 11 specimens, including partial bones from the hand, foot, arm, and clavicle, were found representing at least five individuals. Like Ar. ramidus, this creature was similar in body and brain size to a chimpanzee, but was claimed to be over a million years older than Ar. ramidus (5–6 million years old).
It comes as no surprise that Ar. ramidus was claimed to be capable of bipedal walking. But the evidence for this is based on a single bone from the big toe found ten miles away from the other fossil specimens! It is claimed that this toe has a broad, robust appearance used for the push-off that is distinctive of the bipedal human gait. This sort of evidence is only convincing to those who begin with the premise that humans have evolved from apelike animals.
Australopithecus Group
Like the Ardipithecus group, the australopithecines are another group of apelike hominins found in eastern Africa, with most found in Hadar in the Afar region of the Great Rift Valley in Ethiopia. These Hadar australopithecines are claimed to range in age from 2 to 4 million years old. Although evolutionists concede that they were clearly adept as tree climbers, the Australopithecus group are claimed to have walked upright on a regular basis. The scientific name Australopithecus means “southern ape,” suggesting, whatever else might be said about these creatures, they are apes. The distinguished paleoanthropologist Lord Solly Zuckerman rejected the notion that australopithecines have anything to do with human evolution, and is reported to have said, “They are just bloody apes.”22
Australopithecus anamensis
At a claimed age of 3.9 to 4.2 million years old, Au. anamensis is believed to be the oldest of the australopithecines, even older than Lucy which Au. anamensis resembles. The first fossil of this type was found in Kanapoi in northern Kenya in 1995 by a team led by Meave Leakey and coworkers. The fossil consisted of several teeth and bone fragments.
But 30 years earlier, in 1965, a single arm bone (distal end of the humerus) known as KNM-KP 271 was discovered at the same site and was dated at 4.5 million years old, making it the oldest human ancestor that had been found at that time. The problem is that studies using computer discriminate analysis have shown that the Kanapoi humerus is strikingly similar to that of modern Homo sapiens. For example, the evolutionist Henry M. McHenry concluded, “The results show that the Kanapoi specimen, which is 4 to 4.5 million years old, is indistinguishable from modern Homo sapiens.”23 The only reason that evolutionists dismiss this humerus as being human is that they consider it to be far too old to be human. If it had been found in strata considered to be less than 200,000 years old, it would be considered to be unquestionably human. As a result, this apparently human humerus is now considered to be Au. afarensis.
Australopithecus africanus
Raymond Dart acquired this very apelike fossilized skull in 1924, and after 20 years of debate it was the first fossil to convince evolutionists that humans had their origin in Africa. Called the “Taung child,” dates proposed for it have ranged between 3.3 and 0.87 million years. Still, it is generally considered the earliest ancestor of humans found in southern Africa. The “Taung child” is a juvenile estimated to be 3 to 4 years of age. Though Dart was convinced of its direct ancestry to man, he published no detailed study of the skull.
Much has been made of the supposed human-like brain development of this creature based on cranial sutures. In humans, the continued rapid growth of the brain after birth is made possible by the skull bones being separated by fibrous connective tissue called the fontanelles. These skull sutures generally close between 12–14 months of age, but may not completely close until about five years old. In apes, with their much smaller brain development, the cranial sutures are closed or nearly closed at the time of birth. Evolutionists have argued that the Taung skull has signs of human-like delayed cranial suture closure. But this has been called into question by a more recent study.24
Australopithecus afarensis
Australopithecus afarensis derives its species name from the Afar region of Ethiopia where most of the apes of this type have been found. Bones and teeth from over 300 individuals of this species have been found and “dated” from 3.85 to 2.95 million years old. The most famous and most complete specimen, popularly known as “Lucy,” has become a commonly used synonym for the whole species Au. afarensis. Lucy has been considered the “gold standard” by which other apelike ancestors of man are judged and is certainly the best known among laymen. For this reason, we will go into greater detail in critically evaluating this fossil.
Lucy was discovered in the Afar region of Ethiopia in 1974 by a team led by the American paleoanthropologist Donald Johanson. Lucy’s fossilized skeleton consists of 47 bones (comprising about 20% of the whole skeleton), including parts of both upper and lower limbs, vertebrae, ribs, and pelvis. While most of the hand and foot bones are missing, as well as most of the skull (except for the mandible and cranial fragments), these have been partially filled in by the discovery of other presumed specimens of Au. afarensis. Lucy appears to have been a fully grown female primate that would have stood about 3.5 feet tall.
Lucy has been widely portrayed in the media and in museums all over the world as the supreme example of a “missing link” between apes and humans, though she is not considered to be directly related to humans. Still, Lucy has been touted as the “mother of all human kind,” and her fossil bones were displayed on a tour of major museums in America under the banner “Lucy’s Legacy—Her Story Is Your Story.” Most creationists consider Lucy and all the other australopithecines to be nothing more than extinct apes with similarities to both chimpanzees and gorillas.
Skull
Even a layman would be unlikely to confuse the skull of Au. afarensis with a human skull. The cranial capacity is small even by ape standards. The sloping face, flat nasal bones, and robust gorilla-like jaw are all typical of ape skulls. But evolutionists claim the teeth of Au. afarensis were human-like with relatively thick enamel and relatively small canine teeth. But as noted earlier, some living apes have these characteristics, so this is not decisive.
Pelvis
A great deal of attention has been given to the pelvic bones of Lucy in an effort to support the possibility of bipedal locomotion. The orientation of the pelvic iliac blades is a key requirement for bipedality. The orientation of the iliac blades on the human pelvis allows the human to use gluteal muscles to counterbalance the lifting of the opposite leg during bipedal walking. This stabilizes the hips and prevents the collapse of the hip on the side of the lifted leg. The orientation of the straight iliac blades of the ape pelvis, in contrast, provides no stability to the hip, forcing the ape to swing the upper body from side to side to keep from falling over sideways with each step.
Evolutionists Stern and Susman25 as well as Tuttle26 believed that Lucy’s pelvis was well-adapted for arboreal (tree-dwelling) life. Like many others, they noted that the orientation of the pelvic iliac blades was similar to that of chimpanzees, not humans. But Owen Lovejoy, who worked with Johanson analyzing the Lucy fossils, believed that the first reconstruction of Lucy’s pelvis was in error and, in a much-publicized video shown on public television, Lovejoy demonstrated how casts of Lucy’s pelvis could be cut apart and rearranged to produce the desired more human-like pelvis that might be capable of human-like bipedal locomotion.27
Other analyses of the pelvis by evolutionist Christine Berge, who has taken advantage of modern technology, offer a different reconstruction.28 Berge writes, “The results clearly indicate that australopithecine bipedalism differs from that of humans.” She pointed out that the extended limb of australopithecines would have lacked stabilization during walking, suggesting a retention of a partly arboreal behavior.
Feet and footprints
Much of the presumed evidence for Lucy walking upright in a human-like bipedal fashion, comes from the evidence of footprints found by Mary Leakey in the Laetoli site in Tanzania. Leaky discovered a 73-foot-long trail of fossilized footprints consisting of 20 prints of an individual the size and shape of a modern ten-year-old human and 27 prints of a smaller person. The paleoanthropologist Timothy White, who was working with Leakey at the time, said,
Make no mistake about it, they are like modern human footprints. If one were left in the sand of a California beach today, and a four-year-old were asked what it was, he would instantly say that somebody had walked there. He wouldn’t be able to tell it from a hundred other prints on the beach, nor would you. The external morphology is the same. There is a well-shaped modern heel with a strong arch and good ball of the foot in front of it. The big toe is straight in line. It doesn’t stick out to the side like an ape toe.29
Louis Robins of the University of North Carolina, who also analyzed the footprints, said, “The arch is raised, the smaller individual had a higher arch than I do—the toes grip the ground like human toes. You do not see this in other animal forms.”30
In a lecture I attended at Washington University in St. Louis in 1982, Mary Leaky pointed out one additional feature of her Laetoli footprints rarely mentioned in the literature—all of the larger footprints of the trail have a smaller footprint superimposed on them! This could be interpreted as a child increasing its stride to step in an elder’s footprints, as children often do. In addition, she reported that there were tracks of a wide variety of animals that are similar or identical to animals living in the area today, including antelopes, hares, giraffes, rhinoceroses, horses, pigs, and two kinds of elephants. Even several bird eggs were found, and many of these could be easily correlated with eggs of living species.
Mary Leaky assumed that the footprints were made by some hominid, but not by Homo sapiens, because the stratum in which the prints are found is estimated to be 3.66 million years old. Since that falls roughly near the presumed age of Au. afarensis, Johanson has steadfastly insisted that they must have been made by Au. afarensis. This conclusion is made despite the fact that the Laetoli footprints are located about 1,000 miles away from the location of the Hadar australopithecines such as Lucy.
Mary Leaky disagreed with Johanson’s claims for Au. afarensis as the maker of her footprints. Also, two groups of scientists working independently have challenged the claim that Lucy had completely abandoned the trees and walked fully upright on the ground.31 Anthropologist Russel Tuttle from the University of Chicago said that the Laetoli footprints that Leaky discovered in Tanzania were made by another more human species of ape-man that coexisted with Au. afarensis about 3.6 million years ago and that it was this unknown hominid that is the direct ancestor to man. After a careful examination of the Laetoli prints and foot bones of the Hadar Au. afarensis Tuttle concluded that the Hadar foot is apelike with curved toes, whereas the footprints left in Laetoli are virtually human.32
Australopithecus sediba
In 2008, the young son of paleoanthropologist Lee Berger found the first specimen named Australopithecus sediba in the Malapa cave north of Johannesburg. The elder Berger, who is well known for self-promotion, reported the discovery to mixed reviews in the April 9, 2010, issue of Science. The species name sediba means “wellspring” and is commonly known as “Malapa boy.” The problem is that with a claimed age of 1.78–1.95 million years old Au. sediba is considered by most evolutionists to be too young to be an ancestor of Homo.
There are many nontrivial differences between Au. sediba and humans. Au. sediba has a brain measuring about one-third the size of that of a typical human but well within the range of apes. A comparison of the skull of Au. sediba with that of humans reveals that the lower face of Au. sediba is sloped like that of apes. And, like apes, the forehead of Au. sediba is flat, making the orbits of the eyes barely visible when viewed from the side. The mandible of Au. sediba bears no close resemblance to that of man (or even a chimpanzee) but rather is more similar to that of a gorilla.
The postcranial skeleton of Au. sediba is also very apelike. It has a small body with apelike large-jointed upper and lower limbs. The arms and hands of Au. sediba extend down to the knees, typical of long-armed knuckle walkers. The up-tilted glenoid fossa (shoulder joint) of Au. sediba, together with its long, curved fingers are typical of the suspensory adapted upper limbs of tree-dwelling apes. The feet are described as primitive and similar to other Australopiths. But are these striking differences between Au. sediba and humans outweighed by the similarities?
Paleoanthropologist Tim White said, “The characteristics shared by A. sediba and Homo are few and could be due to normal variation among australopithecines.”33 The claimed “Homo-like” features of the Au. sediba pelvis are based on a composite reconstruction of the juvenile MH1 specimen. Any claim of human-like bipedality based on this pelvis is necessarily speculative and subject to the bias of the observer. Au. sediba is claimed to have protruding nasal bones like that of humans (and unlike the flat nasal bones of living apes). But this “similarity” is based on a slight bony raphe between otherwise flat nasal bones typical of australopithecines in general.
Paranthropus Group
The members of this group were formerly considered to be a more robust type of australopithecine, but now have been given the genus name Paranthropus. All are very apelike with large teeth, massive jaws, and large chewing muscles. These apes are claimed to have lived 1.2 to 2.7 million years ago, making them broadly overlap in time with the australopithecine group. There is no compelling reason to think that any of these apes had anything to do with humans. But evolutionists consider the members of this group to be bipedal based on perceived similarities in the hip joint and big toe to that of humans.
Paranthropus robustus
In 1938, Robert Broom discovered a very apelike fossil jaw fragment and molar in Kromdraai, South Africa. Other teeth and bones were found later and attributed to the genus Paranthropus, which fancifully means “beside man.” This ape had a highly sloped face and a large gorilla-like sagittal crest on its skull for the attachment of massive chewing muscles. About the only thing that can be said to be human-like about this ape is relatively thick tooth enamel.
Paranthropus boisei
In 1959, Mary Leakey discovered the most famous fossil specimen of this species, commonly known as “Zinj” (short for Zinjanthropus), in the Olduvai Gorge of northern Tanzania. It is very similar to the other members of the Paranthropus group in being very apelike with a broad sloping face, gorilla-like sagittal crest, massive chewing muscles, and small brain. The cheek teeth were even larger than those of P. robustus and about four times the size of human molars. Once again, the relative thickness of the tooth enamel is sighted as being “human-like,” but that again is not a uniquely human quality.
Paranthropus aethiopicus
Evolutionists debate whether or not this is really a different species from the other robust australopithecine, paranthropithecine, and zinjanthropithecine apes. One problem is the scanty fossil evidence for this species, which consisted only of a toothless partial mandible until Alan Walker and Richard
Leakey found a skull (known as the “Black Skull”) west of Lake Turkana in eastern Africa that they believed to be of the same species. Like the other Paranthropus fossils, P. aethiopicus is very apelike with the usual sagittal crest, massive jaws and teeth, huge chewing muscles, and sloping face.
Homo Group
With the Homo group, excepting Homo habilis, we suddenly encounter humans. Not only are they large brained and anatomically human with unambiguous evidence of human-like bipedality, but we also find evidence of tools and other artifacts unique to humans. It should be noted that evolutionists believe that the Homo group dates from the present back to 2.4 million years ago, making them broadly overlap with Paranthropus group, which in turn broadly overlap with the Australopithecus group. This means that when we find evidence for tools, butchery, and use of fire, we cannot reflexively attribute them to the nonhuman genera of hominins. If a modern land-filled garbage dump were excavated, we might find chicken bones in close association with plastic spoons and forks, but this is hardly evidence that the chicken was the “tool maker.”
Homo habilis
Homo habilis is considered to be one of the earliest members of the genus Homo that are presumed to have lived between 1.4 and 2.4 million years ago. But even many evolutionists consider H. habilis to be an empty taxon consisting of a collection of several dozen controversial and confusing fossil specimens.
It all began in 1964 when Louis Leaky, Phillip Tobias, and John Napier reported in Nature the discovery of fossil skull fragments as well as hand and foot bones from four juvenile specimens. The foot and hand bones were reported to be human-like and capable of human-like functionality, but the skull was more apelike. But the skull bone fragments were not found next to the foot and hand bones, leading some scientists to conclude that the H. habilis fossils were actually a mixture of H. erectus and australopithecine fossils.
Some years earlier, Louis Leaky found stone tools at the same site and attributed them to a fossil he called Zinjanthropus, now considered to be an ape. With the discovery of the new fossils, Leakey declared them to be the toolmaker, thus the name Homo habilis, which means “handy man.”
To add to the confusion, in 1986, Tim White and Donald Johanson discovered fossil bones in Olduvai Gorge, which were “dated” to be 1.8 million years old and also claimed to be H. habilis on the basis of similarities of the skull and teeth. But this fossil, known as Olduvai Hominid 62, had an apelike body that was even smaller than the australopithecine Lucy, which was only a little over three feet tall. Indeed, some evolutionists regard H. habilis to be australopithecine.
Homo rudolfensis
The first, and some would say the only, specimen of Homo rudolfensis is a skull discovered in 1986 by Richard Leaky in Koobi Fora in the Lake Turkana basin of Kenya. It is perhaps best known by Leakey’s original designation, KNM-ER 1470. First “dated” at three million years old, it is now dated at 1.9 million years (based on pig fossils found nearby34) in an effort to make it more consistent with its human-like appearance.
Skull 1470 serves as a good example of the problems encountered in interpreting the reconstruction of highly fragmented fossils. The nearly 300 fragments required two months to reconstruct into a remarkably complete skull. The cranial capacity was estimated to be nearly 800 cm3. Considering the relatively small size of the skull, this is well within the range of modern human variability. But the relatively long sloping face below the orbits gives 1470 an apelike appearance. The problem is the maxilla (upper jaw) of this specimen is attached to the cranium by only a narrow shaft of bone, allowing the face to look sloped like an ape or vertical like a human. Alan Walker, who worked with Leaky in interpreting 1470, said, “You could hold the maxilla forward and give it a long face, or you could tuck it in making the face short. How you held it really depended on your preconceptions.”35 All commercially available replicas of the 1470 skull show the maxilla swung forward giving an apelike appearance.
If one ignores the presumed “date” of 1470 and examines the skull with maxilla swung in, it has the distinct appearance of a human skull. The orbits are slightly wider than tall, and are visible when viewed from the side, much like a human skull. There is no trace of a brow ridge or sagittal crest. The forehead is tall and nearly vertical as in humans. If this skull had been found in strata believed to be less than 200,000 years old, it would have been declared to be a human.
Homo erectus
Homo erectus includes the early fossil skull “Java man” discovered by the Dutch anatomist Eugene Dubois in 1891, and “Peking man” found in Zhoukoudian, China, which comprised 200 human fossils from more than 40 individuals, nearly all of which were lost during World War II. Evolutionists believe H. erectus lived between 1.9 million and 144,000 years ago, overlapping with the Paranthropus group and modern Homo sapiens.
The most complete fossil considered to be H. erectus is known as “Turkana Boy,” discovered by Richard Leaky in 1984 near Lake Turkana in Kenya. This nearly complete skeleton consisting of 108 bones has been estimated to be about 11–12 years old. The pelvic anatomy indicates that this individual would have been fully bipedal. The brain size was estimated to be about 800 cm3. Unlike apes, Turkana Boy shows clear evidence of protruding nasal bones.
In general, H. erectus had human-like body proportions with relatively short arms and long legs compared to the size of the torso. These fossils are often found in association with tools such as hand axes and cleavers, as well as evidence of fire hearths for cooking.
Homo heidelbergensis
The first fossil specimen of this type was found in 1908 near Heidelberg, Germany. The specimen was a complete mandible missing the premolars and first two left molars. Most of the fossil specimens claimed to be of this type have been found since 1997 in northern Spain, Ethiopia, Namibia, and South Africa. It is claimed H. heidelbergensis lived between 700,000 and 200,000 years ago.
There is an absence of any clear dividing lines between H. erectus, H. heidelbergensis, and Neanderthals. As a result, some evolutionists classify H. heidelbergensis as H. erectus. The problem is that there are no obvious transitions among this group and no unique characteristics that clearly distinguish H. heidelbergensis from H. erectus and Neanderthalensis. This chaos in classification serves to emphasize that evolutionists are not even in agreement on what exactly constitutes the genus Homo.
Fossil evidence indicates that the male H. heidelbergensis averaged about 5 feet 9 inches tall with a cranial volume of 1100–1400 cm3, well within the range of normal modern human variability. There is evidence H. heidelbergensis built shelters, buried their dead, and made hand axes and stone-tipped spears for hunting and various other wooden tools.
Homo neanderthalensis
As Lubenow will discuss more fully in the next chapter, the fossilized bones of nearly 500 Neanderthals have been found since the first recognized.
Neanderthal fossils were discovered in the Neander Valley near Dusseldorf, Germany, in 1856. Neanderthal fossils have been found throughout most of central and western Europe, the Carpathians, Balkans, Ukraine, western Russia, and northern Asia. So far, none have been found in Africa. Evolutionists claim that Neanderthals lived between 400,000 and 40,000 years ago, making them broadly overlap with modern Homo sapiens.
Neanderthals are clearly human in every respect, having no physical traits that fall outside the range of normal human variability. But the cultural evidence makes their fully human nature even more compelling. Neanderthal fossils have been found in association with numerous artifacts such as tools, musical instruments, jewelry, cooking hearths, fabricated shelters, and evidence that they buried their dead. For many years, Neanderthals have been classified as Homo sapiens with a subspecies name neanderthalensis. More recently, the name has been changed to Homo neanderthalensis, in an effort to distinguish the Neanderthals from modern man.
Homo floresiensis
The fossils designated Homo floresiensis were first found in 2003 on the Island of Flores in Indonesia, and thus far they have been found nowhere else. These fossils are commonly referred to as “Hobbits” because they are estimated to have been only about 3 feet 6 inches tall! To make matters even more enigmatic, bones throughout their skeletons were deformed in places. Evolutionists have been in considerable disagreement about the correct interpretation of the fossil evidence, but the evidence indicates that they were most likely true humans who remained small and deformed because of disease (such as microcephaly or Down syndrome) or malnutrition. They certainly present no evidence for human evolution from apes.36
Homo sapiens
Evolutionists believe that Homo sapiens (anatomically modern humans) evolved in Africa about 200,000 years ago. The main difference between H. sapiens and most other members of the Homo group is that H. sapiens is less robustly built. Differences in cranial capacity, pronounced brow ridges, and jaw anatomy are inconsistent.
Fossils of several anatomically modern humans have been found, including the following:
- Cro-Magnon man—Europe
- Neanderthals—western Europe and Asia
- Grimaldi man—Italy
- Chancelade man—France
- Predmosti—Czech Republic
- Denisovan—southwestern Siberia
There is no agreement among evolutionists regarding which if any of the other claimed species of Homo is our direct ancestor. Indeed, not a single hominin can be shown to be the direct ancestor of any other; all is conjecture. But evolutionists generally maintain that Homo evolved from some australopithecine.
Conclusion
As we have seen in this survey of the “evidence” presented in the traveling exhibit of the Smithsonian Museum (in a public library near you some day), the classification of man’s presumed ancestors (hominins) is constantly in flux and is a subject of often testy disagreements among paleoanthropologists themselves. In contrast, the modern classification of living humans is very straightforward, leaving no room for confusion. Thanks to the success in sequencing the human genome, there is now a consensus among scientists that there is only one living genus of humans (Homo), one species (sapiens), one subspecies (sapiens), and one race (human). This is certainly consistent with all humans sharing ancestry with one pair of human parents, Adam and Eve. There is no scientific reason to doubt the literal truth of Genesis 1–5 about their origin (or the origin of anything else in those chapters).
Searching for Adam

You can believe what the Bible says about Adam and man's origin and with intellectual integrity reject the myth of human evolution.
Read Online Buy Book
Master Books has graciously granted AiG permission to publish selected chapters of this book online. To purchase a copy please visit our online store.
Footnotes
- Charles Darwin, https://www.darwinproject.ac.uk/letter/DCP-LETT-2192.xml.
- Charles Darwin, The Origin of Species by Means of Natural Selection (London, England: Penguin Books, 1985, reprint of 1859 1st ed.), p. 205.
- Andrew Snelling, “Where Are All the Human Fossils?” Creation Ex Nihilo 14:1 (1991): 28–33.
- Kurt Wise, “Completeness of the Fossil Record,” https://answersingenesis.org/fossils/fossil-record/completeness-of-the-fossil-record/, November 23, 2009.
- Michael Denton, Evolution: A Theory in Crisis (London, England: Burnett Books, 1985), p. 190.
- Bjorn Kurten, Pleistocene Mammals of Europe (Chicago, IL: Aldine, 1968).
- http://humanorigins.si.edu/evidence/human-fossils.
- Julien Huxley, “Evolution and Genetics,” in What Is Science, ed. J.R. Newman (New York, NY: Simon and Schuster, 1955), p. 278.
- Antony Bushfield, “Chimps Believe in God, Research Claims,” https://www.premier.org.uk/News/World/Chimps-believe-in-God-researcher-claims, March 4, 2016.
- Canadian Institute for Advanced Research, “Human-like ‘Eye’ in Single-celled Plankton: Mitochondria, Plastids Evolved Together,” ScienceDaily, www.sciencedaily.com/releases/2015/07/150701133348.htm, July 1, 2015.
- Anon., “The Foramen Magnum: How Do We Know?” https://afarensis99.wordpress.com/2006/04/22/the_foramen_magnum_how_do_we_k/, April 22, 2006.
- J.C. Ahern, “Foramen Magnum Position Variation in Pan troglodytes, Plio-Pleistocene Hominids, and Recent Homo sapiens: Implications for Recognizing the Earliest Hominids,” American Journal of Physical Anthropology 127:3 (2005): 267–276.
- “A Discovery of Supreme Importance to All Interested in the History of the Human Race,” Illustrated London News, December 28, 1912, p. 4–5.
- Glen Levy, “Piltdown Man,” TIME, http://content.time.com/time/specials/packages/article/0,28804,1931133_1931132_1931125,00.html, March 16, 2010. Levy says that no culprits other than Dawson have ever been implicated. But actually, evidence points to the involvement of some of the leading British scientists at the time. See Robin McKie, “Piltdown Man: British Archeaology’s Greatest Hoax,” https://www.theguardian.com/science/2012/feb/05/piltdown-man-archaeologys-greatest-hoax, February 5, 2012.
- Levy, ibid.
- Solly Zuckerman, Beyond the Ivory Tower (New York, NY: Taplinger Publishing Co., 1970), p. 65.
- F. Clark Howell, Early Man (New York, NY: Time-Life Books, 1965), p. 41–45.
- Bernard Wood, “Who are we?” New Scientist, 2366 (October 26, 2002): 44.
- For creationist historical analysis of this discovery with the LIN picture, see Ian Taylor, In the Minds of Men 6th ed. (Foley, MN: TFE Publishing, 2008), p. 227–229. The five-year process of reinterpreting the fossil evidence is described by William K. Gregory, “Hesperopithecus Apparently Not An Ape Nor A Man,” Science, 66:1720 (December 16, 1927): 579–581.
- http://humanorigins.si.edu/evidence/human-family-tree.
- The abstract to the PNAS technical article is linked in Sergio Prostak, Natali Anderson, and Enrico de Lazaro, “Study: Last Common Ancestor of Humans and Apes Looked Like Gorilla or Chimpanzee,” http://www.sci-news.com/othersciences/anthropology/science-homo-pan-last-common-ancestor-03220.html, September 9, 2015.
- Quoted in Roger Lewin, Bones of Contention (New York, NY: Touchstone Books, 1988), p. 165.
- Henry M. McHenry, “Fossils and the Mosaic Nature of Human Evolution,” Science 190:4213 (1975): 425–431.
- R.L. Holloway et al, “New High-resolution Computed Tomography Data of the Taung Partial Cranium and Endocast and Their Bearing on Metopism and Hominin Brain Evolution,” Proceedings of the National Academy of Sciences USA 111:36 (2014): 13022–13027.
- J.T. Stern and R.L. Susman, “The Locomotor Anatomy of Australopithecus afarensis,” American Journal of Physical Anthropology 60 (1983): 279–217.
- R.H. Tuttle, “Knuckle-walking and the Problem of Human Origins,” Science 166:3908 (1969): 953–961.
- Donald Johanson, “In Search of Human Origins (Part 1),” Nova, PBS Airdate: June 3, 1997. Transcript at http://www.pbs.org/wgbh/nova/transcripts/2106hum1.html.
- Christine Berge and Dionysis Goularas, “A New Reconstruction of Sts 14 Pelvis (Australopithecus africanus) from Computed Tomography and Three-dimensional Modeling Techniques,” Journal of Human Evolution 58 (2010): 262–272.
- Donald Johanson and Maitland Edey, Lucy, The Beginnings of Humankind (New York, NY: Simon and Schuster, 1981), p. 250.
- Louis Robins, quoted in Anon, “The Leaky Footprints: An Uncertain Path,” Science News 115:13 (March 31, 1979): 196.
- Anon., “Was Lucy a Climber?” Science News 122:8 (1982), p. 116.
- R.H. Tuttle, D.M. Webb, and M. Baksh, “Laetoli Toes and Australopithcus Aferensis,” Human Evolution 6:3 (1991): 193–200.
- M. Balter, “Candidate Human Ancestor From South Africa Sparks Praise and Debate,” Science 328:5975 (2010): 154–155.
- The fascinating account of the ten-year dating controversy is reported in Marvin Lubenow, Bones of Contention (Grand Rapids, MI: Baker, 1992), p. 247–266.
- Quoted in Roger Lewin, Bones of Contention (New York, NY: Touchstone Books, 1988), p. 160.
- Elizabeth Mitchell, “Is Homo floresiensis a Legitimate Human ‘Hobbit’ Species?” https://answersingenesis.org/human-evolution/homo-floresiensis/homo-floresiensis-legitimate-human-hobbit-species/, August 19, 2014.




Support the creation/gospel message by donating or getting involved!

Answers in Genesis is an apologetics ministry, dedicated to helping Christians defend their faith and proclaim the good news of Jesus Christ.
- Customer Service 800.778.3390
- © 2024 Answers in Genesis